

Семейство: Фазановые Phasianidae
Род: Улары Tetraogallus
Вид: Tetraogallus himalayensis (Gray, GR 1843)
Протоним: Tetraogallus Himalayensis.
Синоним: Tetraogallus himalayensis, Tetraogallus himalayensis himalayensis, Tetraogallus nigellii.
Немецкое название: Himalajakönigshuhn
Испанское название: Perdigallo Himalayo, Tetraogallo del Himalaya.
Казахское название: гималай күркетауығы, Гималай ұлары, гималайлық ұлар, қарабауыр ұлар
Украинское название: Улар гімалайський.
Другие названия: темнобрюхий улар, гималайская горная индейка.
Taxonomic Serial Number : 175953 | Global Biodiversity Information Facility : 2473455.
Список ПОДВИДОВ вида Гималайский улар Tetraogallus himalayensis (Gray, GR 1843)
‐ . — Tetraogallus himalayensis sauricus (Potapov, RL 1993)
‐ . — Tetraogallus himalayensis sewerzowi (Zarudny, NA 1910)
‐ . — Tetraogallus himalayensis incognitus (Zarudny, NA 1911)
‐ Туркестанский темнобрюхий улар — Tetraogallus himalayensis himalayensis (Gray, GR 1843)
‐ Кашгарский темнобрюхий улар — Tetraogallus himalayensis grombczewskii (Bianchi, VL 1898)
‐ Наншанский улар — Tetraogallus himalayensis koslowi (Bianchi, VL 1898)
ОСНОВНОЕ ОПИСАНИЕ ПО: Гималайский улар — Tetraogallus himalayensis (Gray, GR 1843)
Окраска. Взрослый самец. Окраска верхней части тела светло-серая, с голубоватым оттенком и желтовато-песочным налетом на передней части спины. На голове — немного светлых пестрин, в области зашейка — ряд коричневых пестрин ошейника, охватывающего основание шеи. У многих особей этот ошейник сплошной, у других — разорван в области зашейка. Лопаточная область самая темная, с шиферным оттенком, причем вся спина, крылья и поясница с мелким струйчатым рисунком. Кроме того, в лопаточной области, на крыльях и пояснице развит продольный рисунок из желтовато-коричневых штрихов, образованных боковыми полями вершин перьев. По бокам головы и шеи две коричневые полосы: одна идет от ушной области к основанию шеи, вторая — от угла рта тоже к основанию шеи, где они соединяются упомянутым выше ошейником. Бока и передняя часть шеи между этими полосами, а также горло белые. Лоб и полоса над глазом беловатые, бока головы голубовато-серые. За глазом небольшое голое пятно, обычно скрытое соседними перьями, но в брачный сезон приобретающее желтый цвет и становящееся хорошо заметным. Грудь белая, отороченная спереди ошейником с правильным поперечным рисунком из черных коротких полос, образованных предвершинными полосами грудных перьев. Брюхо темное, серо-бурое, с тонким струйчатым рисунком. По бокам тела идут широкие продольные полосы черного цвета с рыжей окантовкой. Первостепенные маховые перья белые с широкими бурыми вершинами, второстепенные маховые сохраняют только белые основания, а серый струйчатый рисунок заходит по краям опахал далеко вниз, вершины их с желтовато-коричневым струйчатым рисунком. Рулевые перья с коричневыми наружными и бурыми внутренними опахалами, на центральных рулевых перьях сплошь развит струйчатый коричневый рисунок, постепенно исчезающий на боковых перьях. Вершины рулевых перьев коричневые, на боковых парах намечается черная предвершинная полоса. Нижние кроющие перья хвоста белые. Клюв серый, ноги красновато-желтые.
Взрослая самка. Окраска такая же, как и у самца, только черные полосы на груди развиты меньше. Признак этот, однако, ненадежный и основным критерием при определении пола взрослой птицы все же являются ее размеры и наличие или отсутствие шпоры.
Самец и самка в ювенальном наряде. Преобладает темно-серый цвет. На верхней части тела (на зашейке и передней части спины) по серому фону разбросаны мелкие желтоватые пятнышки, задняя часть спины и поясница в тонких черных поперечных полосках. Верхняя часть головы серовато-голубоватая, коричневый ошейник в основании шеи только намечен, Над глазом белая бровь, окаймленная по сторонам тонкими черными полосками, еще одна темная полоса намечается от угла рта к основанию шеи. На кроющих перьях крыла и на внутренних второстепенных маховых ланцетовидный рисунок из струйчатых коричневых и бурых полос, вытянутых вдоль опахал, остальные маховые перья бурые, со струйчатым рисунком по краю внешнего опахала. Окраска нижней части тела светлее: горло беловатое, коричневато-серая грудь покрыта мелкими белесыми пятнышками, низ груди и брюхо серые, с неясным поперечным рисунком из светлых полосок на передней части брюха.
Пуховой наряд. Пуховой птенец темно-серый сверху, со сложным узором из темных и светлых полос. На голове сложный рисунок из черно-бурых полос и пятен. С каждой стороны головы выделяются 4 темные продольные полосы: по бокам темени, над и под глазом (спереди они сливаются) и от угла рта к основанию шеи. Низ тела белесый с серым налетом на груди.
Строение и размеры. Крылья короткие и тупые, но вершина крыла сдвинута кнаружи и приходится на 9-е и 8-е первостепенные маховые. Хвост короткий, закругленный. Ноги высокие, сильные, на плюсне самцов толстая шпора, длиной 10—12 мм. У самок вместо нее имеется только небольшой бугорок. Клюв сильный, слегка загнутый вниз.
Самцы (п = 14, кол. ЗИН АН СССР):
крыло 326.9 (304—356),
хвост 202.8 (195—209),
длина клюва 26.7 (24.7—29.6),
ширина клюва 16.6 (15.7—18.8),
плюсна 68.9 (60—72.3),
средний палец без когтя 56.5 (51.8—60.1).
Самки (п=9, кол. ЗИН АН СССР):
крыло 283.4 (283—310),
хвост 177.1 (163—192),
длина клюва 25.3 (22.9—26.4),
ширина клюва 15.4 (13.8—17.1),
плюсна 61.1 (55.3—65.4),
средний палец 52.5 (51—55.4).
Сезонная динамика массы не изучена. Масса самцов колеблется в пределах 2300—3375, самок 1750—2900. Масса птиц максимальна в ноябре и минимальна в конце мая—начале июня. Молодые птицы в первую осень достигают массы 1800—1900 (Кузьмина, 1962).
Подвиды. Номинативный подвид, населяющий Гималаи, Гиндукуш и западную часть Памиро-Алая, отличается темной окраской, особенно черноватым налетом на груди и почти полным отсутствием коричнево-желтых тонов. Ошейник, как правило, замыкается на зашейке.
Второй подвид, Т.h. sewerzowi Zarudny, 1910, населяющий северную часть ареала (Тянь-Шань, Заилийский Алатау, Тарбагатай, Саур) отличается коричневым оттенком окраски; ошейник на зашейке замкнут далеко не всегда.
Куньлунь и восточную окраину Памира населяет очень бледко окрашенный Т.h. grombczewskii Bianchi, 1907, у которого зашеек, как правило, одного цвета с головой.
Подвид Т.h. kozlowi Bianchi, 1898, населяющий северо-восточный Тибет с окружающими его горными системами и выделявшийся некоторыми авторами в качестве самостоятельного вида, довольно хорошо отличается от остальных подвидов желтовато-коричневой, а не белой окраской груди.
Суточная активность, социальное поведение. Как уже говорилось, суточная активность уларов стандартна во все времена года и несколько нарушается только в период спаривания и при насиживании кладок. Птицы выходят на кормежку с восходом солнца, кормятся до полудня, затем отдыхают всю середину дня и возобновляют кормежку к вечеру, после чего уходят на ночевку.
Улары — стайные птицы и вне времени размножения держатся обычно небольшими стайками.
Ночевки в летнее время располагаются выше мест кормежек, в закрытых местах среди скал или крупных каменных обломков. О зимних ночевках уже говорилось выше.
Враги, неблагоприятные факторы. Как и для других фазановых птиц, наиболее неблагоприятны для гималайских уларов многоснежные зимы, но ввиду труднодоступности высокогорий для наблюдений при большом количестве снега конкретных сведений по этому вопросу практически нет.
В отношении врагов конкретных данных нет. Взрослые птицы прекращают перекличку и затаиваются, когда в непосредственной близости от них пролетает беркут или бородач. В горах Казахстана неоднократно находили остатки растерзанных уларов, но чьей добычей они были — осталось не выясненным (Кузьмина, 1962).
Хозяйственное значение, охрана. Гималайский улар до сих пор сохраняет свою численность на уровне, не внушающем особого беспокойства за его судьбу. Обитание в труднодоступных горах сохраняет его пока что лучше любого законодательства. Промыслового значения не имел и не имеет, являясь в ряде мест только объектом спортивной охоты. Перспективными представляются также опыты по одомашниванию и вольерному содержанию.этой птицы (Потапов, 1966).
Вопрос об охране гималайского улара может возникнуть только на севере ареала,; где этот вид обитает на сравнительно небольших высотах, причем в;торах, которые все в большей степени подвергаются, хозяйственному освоению человеком.
Южная граница в пределах Тибета не выяснена. Подробности распространения в СССР таковы. Этот вид обитает по всем хребтам Памиро-Алая, превышающим высоту 3000 м над ур. м. Самая западная точка здесь — западная оконечность гор Байсунтау (бассейн Шерабаддарьи), кишлак Земон южнее Пенджикента и вершина 3194 м в западной оконечности Туркестанского хребта (Иванов, 1969; наши данные). По системе Тянь-Шаня он идет на запад до Кураминского, Чаткальского, Пскемского и Угамского хребтов. Населяет Джунгарский Алатау (через хр. Борохоро этот участок связан с тянь-шаньским участком ареала) и, наконец, изолированный участок ареала расположен в хребтах Тарбагатая и Саура (рис. 14).
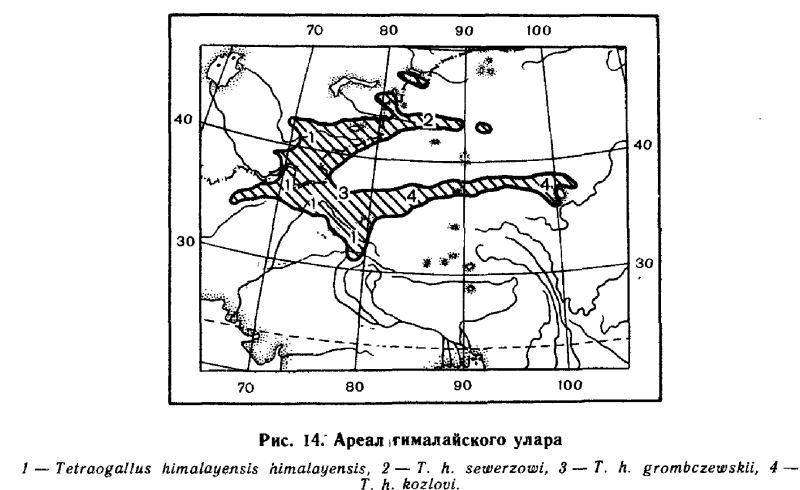
Численность. Данных очень мало. Плотность птиц в горах определять вообще очень трудно из-за крайней неравномерности нх распределения. Отдельные микропопуляции населяют сравнительно небольшие участки, наиболее удобные для жизни, между которыми подчас располагаются обширные пространства, где уларов нет вовсе. На Памире плотность населения в таких удобных местах составляет около 7.5 особей на 100 га весной и 17—20 на ту же площадь осенью (наши данные). Сходные цифры имеются и для Бадахшана: в долине р. Гунт максимальная плотность составляла весной 8—10 птиц (Абдусалямов, 1971). Данные по сезонной и многолетней динамике численности отсутствуют, но в целом в большинстве названных горных районов гималайский улар является вполне обычной птицей.
Наши наблюдения на Памире позволяют считать этим пределом 4800 м над ур.м. Для Гиссаро-Каратегина верхний предел гнездования обусловливается ,как, видимо, и во многих других местах, линией вечных снегов, проходящей здесь на высоте 4000 м (Попов, 1959).
Помимо столь обычных для куриных птиц гастролитов, у зимних птиц в зобах находили и почву, видимо используемую, как источник для получения солей (Кузьмина, 1962). Весной сразу же с началом вегетации улары переходят на свежую зелень — вегетативные и генеративные побеги различных высокогорных трав и полукустарников. Рацион в это время включает цветы, побеги и листья эфемеров — ветреницы, прострела, а также стебли лука. Для Бадахшана, кроме многочисленных эфемеров, в весеннем питании указаны также колоски и стебли костра кровельного, костра острозубого и в небольшом количестве мох, луковицы тюльпанов, листочки иргая и зверобоя (Попов, 1959).
В летнем рационе также продолжает преобладать свежая зелень многих (свыше 100 видов) растений — пырея, мятлика, типчака, астрагалов, лодочника, клевера, купальниц, лютиков, а со второй половины лета семена злаков, бобовых, гречишных, луковицы крокусов и корневища различных растений. Осенью состав пищи тот же, но в рационе Постепенно уменьшается доля зеленых частей растений, пока полностью не начнет преобладать «сухой» корм. В ряде мест отмечена большая доля семян.
Например, в Гиссаро-Каратегине в сентябре зобы птиц были наполнены семенами, колосьями и листьями ястребинок, борца, различных злаков (трещетника, типчака, тимофеевки, мятлика), а также ясколки и мака, причем преобладали семена ястребинок, борца, мятлика и мака (Попов, 1959). Животные корма потребляются летом в очень малых количествах — собственно, это всегда одиночные особи некоторых насекомых, и похоже, что они случайно захватываются с растительным кормом.
В питании птенцов гималайского улара, как и у других видов этого рода, примечательно полное отсутствие животной пищи в первые дни жизни. Позднее, в возрасте 7 дней и старше, они часто поедают отдельных жуков, преимущественно слоников, дождевых червей, но основой постоянно остается растительная пища. Основной же корм в первые дни жизни — нежные листочки различных бобовых растений. Только в редких случаях, при обилии некоторых видов насекомых, животный компонент суточного рациона может быть значительным. Такие случаи известны для Тянь-Шаня, а в. Заилийском Алатау однажды в течение 2 недель улары в большом количестве поедали прямокрылых, и помет их в это время содержал много хитина (Кузьмина, 1977).
Весеннее возбуждение в Бадахшане начинает проявляться уже в начале марта (Абдусалямов, 1971). Громкие мелодичные свисты самцов раздаются все чаще, на рассветах птицы ведут себя все более шумливо и оживленно, и из зимних стай начинают обосабливаться отдельные пары.
В период образования пар и выбора гнездовых участков, судя по отдельным наблюдениям, между самцами происходят ожесточенные стычки: на Памире добывались даже птицы с свежими ранами (Абдусалямов, 1971). В других районах (Заилийский Алатау) драк вообще не наблюдалось (Кузьмина, 1962). Мои наблюдения на Памире начинались с начала мая, когда все улары (за исключением молодых холостых птиц) уже были в отдельных парах, держались у своих участков и драк между ними не происходило. Гнездовые участки отдельных пар, видимо, имеют определенные границы, но каких-нибудь стычек или конфронтаций между самцами в период насиживания на границах участков никто не наблюдал.
Одна из гнездовых территорий, обследованная мной на Памире, начиналась у стен крупного скального массива, где на одном из карнизов располагалось гнездо, и занимала прилегающий снизу склон примерно на 150—200 м вниз, имея в ширину около 100 м. Две соседние пары гнездились в 300—400 м по обеим сторонам от найденного гнезда.
Токовой ритуал самца очень прост. Основной вокальный элемент тока — описанный выше характерный свист. По наблюдениям А.В. Попова (цит. по: Абдусалямов, 1971), свистящий улар «сначала поднимает голову вверх и вперед, в середине свиста — вверх и с окончанием свиста запрокидывает ее назад». Сопровождая кормящуюся самку, самец держит свой хвост почти все время раскрытым, и на темном фоне рулевых перьев резко выделяется белое подхвостье.
Гнездо располагается обычно на склонах южных экспозиций под прикрытием камня, кустика или же в скальной нише. Нередко оно бывает открыто сверху, но большей частью над ним нависает либо каменный выступ, либо ветки кустарника, арчи или ели. Располагается оно вблизи мест, удобных для кормежки, и на Памире, например, большей частью устраивается там, где склоны средней крутизны упираются в скалы.
Выстилка очень скудная, из сухих стеблей и листьев, с небольшим количеством перьев самой наседки. Диаметр гнезда 30—32 см, диаметр лотка 23 см, его глубина около 8 см. Яйца крупные, с гладкой блестящей скорлупой, окрашены в серовато-желтоватый цвет с разбросанными по нему темно-коричневыми мелкими точками и пятнышками. Встречаются яйца почти без пятен.
Масса свежих яиц варьирует от 70 до 89 (Кузьмина, 1962).
Число яиц в кладке колеблется от 5 до 16, в большинстве случаев бывает 6—9 яиц.
В зависимости от места расположения гнезда выводок после вылупления или поднимается наверх, на альпийские лужайки с густой травой, либо же спускается вниз, к верхней границе древесно-кустарниковой растительности. Сроки размножения колеблются довольно сильно, в зависимости от высоты местности и ее географической широты. Наиболее ранняя дата встречи пуховых птенцов у Кульджи — 13 мая (Шестоперов, 1929), самый поздний срок — 3 июля в хр. Кетмень (Кузьмина, 1962).
Сроки размножения могут быть довольно растянуты даже в одной популяции, так что в один и тот же день можно встретить и пуховых птенцов, и лётных молодых птиц. Все же на меньших высотах вылупление птенцов происходит заметно раньше, чем в высокогорье, и птенцы, соответственно, имеют в своем распоряжении гораздо больше времени для развития.
Наиболее поздние сроки размножения в пределах СССР — на Памирском нагорье, где массовое вылупление птенцов происходит в начале июля (Потапов, 1966).
О лётных способностях молодых птиц известно мало. Будучи размерами с перепела, птенцы уже свободно перепархивают на короткие расстояния, а достигнув роста кеклика, они при тревоге взлетают и, как взрослые птицы, улетают вниз по склонам. Видимо, в это время они еще способны взлетать с ровной поверхности и лететь активным полетом.
Участие самца в вождении птенцов несомненно, но далеко не во всех случаях. В процессе насиживания он держится обычно в районе гнезда и всегда сопровождает самку, когда та покидает гнездо и кормится. После вылупления птенцов самцы либо покидают участок и уходят на линьку, присоединяясь к группам холостых линяющих птиц, либо остаются при выводке, который сопровождают вместе с самкой. Если птенцы еще небольшие и не умеют летать, то при появлении опасности самка проявляет очень сильное беспокойство и в большинстве случаев старается отводить от птенцов, притворяясь раненой и издавая квохчущие и стонущие звуки, а после того, как птенцы попрячутся, удаляется на некоторое расстояние и с удобного места наблюдает за нарушителем спокойствия, время от времени издавая такой же свист, как и самцы.
Подрастающие птенцы вместе с самкой, как и остальные взрослые птицы, с приближением осени постепенно поднимаются в своих кочевках на все большие и большие высоты вслед за смещением зоны массовой вегетации и плодоношения растений. В конце августа все птицы, как молодые, так и взрослые, держатся на наибольших высотах, но уже в сентябре начинается их постепенное движение обратно вниз.
При обычном течении жизни улары с рассветом летят описанным выше способом вниз, на кормежки, после чего постепенно поднимаются в процессе кормежки вверх, к местам ночевок, которые достигаются к вечеру.
Во время скользяще-планирующего полета улара слышен громкий свистящий шум. Вверх по склонам улары передвигаются легко и без особых затруднений взбираются по крутым скалам, используя малейшие выступы и карнизы.
Голосовые реакции довольно разнообразны. Наиболее обычная позывка самцов — протяжный свист, начинающийся с низкой ноты и быстро повышающий тональность к концу. К этому свисту прибавляются различные концовки — он или вновь заканчивается низкой нотой, или после него раздается отрывистое квохтанье, или же свист заканчивается низким урчащим звуком, переходящим в отрывистое «ко-ко-ко-ко» с понижением тона в конце. При полетах издается звонкое прерывистое квохтанье, что-то вроде «кли-кли-кли-кли-кулигалу. . .кли-кли-кли-кли-кулигалу-кулигалу» — все эти голоса можно слышать круглый год, но особенно часто и громко они издаются в брачный сезон. Протяжный свист, который многими авторами принимается за брачную вокализацию, на самом деле полифункционален и издается как при тревоге, так и как сигнал своего местонахождения при своеобразных перекличках, происходящих наиболее активно на зорях, но имеющих место и при дневном отдыхе.
Описываются также звуки, напоминающие «лай собаки или плач ребенка» (Кузьмина, 1962). Голосовые реакции самок, видимо, не столь разнообразны. Они более молчаливы, но могут свистеть, как и самцы. При выводках издают негромкое квохтанье.
Улары отличаются очень большой осторожностью. Обладая великолепным зрением и хорошим слухом, они издали замечают опасность и сравнительно легко ее избегают. Их покровительственная окраска делает птиц мало заметными на фоне серых тонов камней и голого грунта.
Крупные размеры, своеобразные полет и голос не позволяют спутать гималайского улара ни с одной другой птицей, и только на Памире, где обитает еще один улар — тибетский, возможна ошибка, особенно на большом расстоянии. Вблизи же гималайский улар легко отличим благодаря темной окраске брюха (белая с пестринами у тибетского улара) и несколько иному голосу.
В Гиссарском хребте улары на зиму спускаются вниз на 1000—1500 м, а при внезапных обильных и длительных снегопадах их встречали на еще меньших высотах. В целом же можно считать, что на большей части ареала улары проводят зиму в пределах древесно-кустарникового пояса растительности, а в некоторых случаях даже выходят на сельскохозяйственные поля предгорий (Шестоперов, 1929). Наряду с вертикальными, происходят и горизонтальные перемещения, но данных о них мало. Видимо, птицы перемещаются также на малоснежные склоны. Незначительной глубины снег уларам не страшен — они легко разрывают его, добывая необходимый корм, но глубокий снег для них губителен. В Бaдaxшaнe, нaпpимep, в прежние времена при глубоких снегопадах местные жители легко добывали уларов, обходя их сверху — увязшие в снегу птицы не могли ни убегать, ни взлетать (Иванов, 1969).
Ночевки птиц всегда происходят в хорошо защищенных от ветра местах — среди скал или в гуще кустарников и даже в пещерах (Корелов, 1956), где, судя по массе помета, птицы ночуют целыми стаями, тесно прижавшись друг к другу. Такие скопления помета уларов встречаются и во многих пещерах Памира. Непосредственных же наблюдений за групповыми ночевками уларов в зимнее время до сих пор нет.
Дневной отдых происходит, как правило, на солнцепеке и энергетически очень важен, поскольку в удачно выбранных безветренных местах даже в высокогорьях Памира воздух и почва днем на солнцепеке нагревается до положительных температур (Потапов, 1966). Кормежки в зимнее время происходят, как и летом, дважды в сутки — утром и вечером, но о степени их длительности данных нет. Сведений о величине суточного рациона также мало. Есть только 2 сообщения о массе сухого содержимого зобов птиц зимой: 42.7 г — 23 декабря на Памире (Абдусалямов, 1971) и 42.5 г—17 февраля в Бадахшане (Попов, 1959). Такое совпадение вряд ли случайно. Если принять во внимание, что улары кормятся дважды в сутки, то суточный рацион, видимо,составляет более 100 г сухой массы.
Всю зиму птицы держатся небольшими стаями по 5—12 особей, обычно не образуя больших скоплений.
В высокогорьях Памира самцы, добываемые в мае, нередко не имеют следов линьки, тогда как на меньших высотах, например в Джунгарском Алатау, они начинают менять оперение уже в конце апреля. Линька на Памире завершается к началу октября. Из особенностей первой дефинитивной линьки следует отметить, что 3 наружных первостепенных маховых (с 8-го по 10-е), вырастающие у птенцов последними, не сменяются, как остальные, а остаются до следующей полной годовой линьки. На наш взгляд, их правильнее рассматривать как дефинитивные, но не имеющие ювенальных предшественников, поскольку их рост начинается уже тогда, когда птица начинает линьку в первый взрослый наряд.
Подробнее о сокращениях можно прочесть здесь.

Песня
…
Позывка
…
Электроманок
…
Литература
• Коблик Е.А. 2025. Фауна птиц стран Северной Евразии в границах бывшего СССР: Списки видов.
• IOC World Bird List v15.1 (Updated February 20, 2025) by Frank Gill, David Donsker & Pamela Rasmussen.
• Н.Н. Балацкий. Птицы Северной Евразии. Таксономический список: август 2023 г.
• «Птицы СССР. Курообразные. Журавлеобразные». т.2. Р.Л. Беме, Н.П. Грачев, Ю.А. Исаков, А.И. Кошелев, E.Н. Курочкин, Р.Л. Потапов, А.К. Рустамов, В.Е. Флинт. Ленинград. Издательство «Наука». 1987 г. стр. 86-95.
Сайты
• zmmu.msu.ru
• www.balatsky.ru
• www.worldbirdnames.org
• Avibase by Denis Lepage
• Zoonomen by Alan Peterson
• Taxonomy by John Boyd
• сайт Василия Вишневского
• www.xeno-canto.org
• www.featherbase.info, www.pbase.com, www.macaulaylibrary.org