

Семейство: Фазановые Phasianidae
Род: Улары Tetraogallus
Вид: Tetraogallus caucasicus (Pallas, PS 1811)
Протоним: Tetrao caucasica.
Синоним: Tetrao caucasica, Tetraogallus caucasicus.
Немецкое название: Kaukasuskönigshuhn
Испанское название: Perdigallo Caucasiano.
Казахское название: Кавказ ұлары
Украинское название: Улар кавказький.
Другие названия: кавказская горная индейка.
Taxonomic Serial Number : 553829 | Global Biodiversity Information Facility : 2473446.
Список ПОДВИДОВ вида Кавказский улар Tetraogallus caucasicus (Pallas, PS 1811)
Подвидов не образует.
ОСНОВНОЕ ОПИСАНИЕ ПО: Кавказский улар — Tetraogallus caucasicus (Pallas, PS 1811)
Окраска. Взрослый самец. Верх темно-серый. Верх головы, уздечка, щеки и задняя часть шеи голубовато-серые, на задней части более или менее выражен рыжеватый налет, который усиливается и становится бурым у старых птиц, особенно на зашейке. Полоса над глазом, кроющие уха, бока и передняя часть шеи до горла включительно белые. Передняя часть спины и верхняя часть груди с поперечно-полосатым рисунком из резко очерченных зигзагообразных черных и светло-охристых полосок. От угла рта или от щек по бокам шеи идут назад к ее основанию широкие полосы черновато-коричневого цвета. Остальная часть спины, поясница, верхние кроющие крыла и хвоста темно-серые с мелким желтоватым струйчатым рисунком и белыми или желтовато-охристыми пятнами по бокам опахал (за исключением кроющих хвоста). На верхних кроющих крыла эти пятна развиты сильнее и имеют более интенсивный рыжий цвет, окаймленный изнутри белым. Маховые перья белые с темно-бурыми вершинами: на сложенном крыле белый цвет их выступает из-под кроющих крыла в виде широкой полосы. Брюхо темно-серое, с мелким охристым крапом и бледно-охристыми окаймлениями опахал. Рулевые перья коричнево-бурые с каштановыми вершинами и узкими белыми вершинными полосками на некоторых перьях (выражены не у всех особей). Бока тела очень темные, в широких продольных полосах коричневого цвета с черными окаймлениями. Нижние кроющие хвоста белые. Оперение ног серое с желтоватым оттенком. Клюв буровато-роговой, ноги оранжево-желтые.
Взрослая самка. Окрашена примерно так же, как и самец, с некоторыми небольшими отличиями. Так, например, полоска, идущая по бокам горла, формируется пестрыми перьями с черно-белым струйчатым рисунком. Перья верха головы и задней части шеи пестрые. Зашеек слабо охристый, не столь ярко окрашенный. Опахала перьев на боках тела имеют более широкие черные и каштановые окаймления, а опахала перьев груди несут более пестрый и яркий рисунок.
Ювенальный наряд (самцы и самки). В общих чертах сходен с окраской взрослых птиц. Лоб, темя, щеки, задняя часть шеи и 2 полосы по ее бокам, идущие от щек, серые с мелким струйчатым рисунком. Горло, шея спереди и 2 полосы от глаз вдоль шеи чисто белого цвета, как у взрослых. Верх груди более светлый и более пестрый, чем ее нижняя часть, и струйчатый рисунок формирует здесь слабо выраженную поперечную полосатость. Нижняя часть груди серого цвета со слабым светло-охристым налетом. Бока тела и брюхо с хорошо выраженным рыжим оттенком, создаваемым светло-охристыми окаймлениями опахал. Подхвостье светло-серое. Весь верх серый, со слабым рыжеватым налетом
и испещрен струйчатым рисунком. Маховые кисти белые, с серыми в пестринах, вершинами. Маховые предплечья серые с беловатым струйчатым крапом. Рулевые серые, с острыми вершинами и охристым струйчатым рисунком по всему опахалу.
Пуховой птенец. Верх темно-серый с многочисленными черноватыми пятнами, образующими сложный рисунок. На голове эти пятна и полоски формируют несколько неясных продольных полос: над и под глазами, по краям темени, по бокам головы от угла рта и назад к затылку. На лбу эти пятнышки сливаются в почти сплошное пятно. Низ тела серый, иногда с слегка желтоватым оттенком. Горло и низ шеи белые. Лапы светлые, клюв рогового цвета со светлой вершиной.
Размеры.
Самцы (п = 2, кол. ЗИН АН СССР):
крыло 280 и 290,
хвост 172 и 190,
длина клюва 23.5 и 25.4,
ширина клюва 15.4 и 16.4,
плюсна 60.7 и 62.1,
средний палец 52.1 и 55.2.
Самки (п = 5, кол. ЗИН АН СССР):
крыло 266 (256—272),
хвост 168.4 (163—176),
длина клюва 22.16 (20.6—24.5),
ширина клюва 14.24 (13.7—15),
плюсна 56.7 (51.8—64.1),
средний палец 50.1 (47.1—52.9).
Базиев (1978) дает следующие размеры.
Самцы (п = 15, декабрь—май):
крыло 277.1 (265—301),
длина тела 584.3 (570—612),
размах крыльев 881.6 (855—935).
Самки (п= 17, тот же период):
крыло 258.7 (245—270),
длина тела 562.7 (540—585),
размах крыльев 827.3 (785—864).
Динамика массы самцов: март—май 1790.7 (1550—1900), июнь—август 1840.3 (1790—2050), сентябрь—ноябрь 2115.4 (1800—2420), декабрь—февраль 1985 (1760—2270). Самки: март—май 1850.8 (1505—2160), июнь—август 1575 (1475—1650), сентябрь—ноябрь
1703.3 (1575—1900), декабрь—февраль 1800.4 (1600—1910).
Суточная активность, социальное поведение. Суточная активность такая же, как и у остальных уларов: ночевка высоко под гребнями гор, спуск утром на кормежку и постепенный подъем в процессе кормежки наверх с отдыхом в середине дня. Только зимой этого дневного отдыха может не быть, и птицы тратят на поиски корма все светлое время суток.
Кавказские улары — стайные птицы и держатся небольшими стайками весь год, исключая только сезон размножения для участвующих в нем птиц.
Враги, неблагоприятные факторы. Врагов у кавказского улара, видимо, немного. Известны случаи преследования уларов орлом могильником, причем без успеха, а также нахождения остатков уларов в зимнем логове рыси (Базиев, 1978). Неблагоприятными для этих птиц являются особенно многоснежные зимы и холодные затяжные весны, приводящие к длительному запаздыванию размножения и прохолостанию многих самок.
Хозяйственное значение, охрана. Наряду с гималайским, кавказский улар пока не внушает особых опасений за свою будущность. Большая трудность охоты на этих птиц и обитание их в местах, где массовый выпас скота затруднен, делает их мало уязвимыми со стороны антропогенных факторов. Общая численность вида оценивается Базиевым в осенний период в 278 тыс. особей, что вполне позволяет использовать этот вид в качестве объекта спортивной охоты в горных охотничьих хозяйствах Главного Кавказского хребта. Однако при этом необходимо проводить такую охоту только в осенние месяцы (октябрь—ноябрь) и только там, где численность птиц не ниже средней, указанной выше. Следует также учитывать высокое эстетическое значение этой птицы, являющейся символом заоблачных высот — ее мелодичный свист, напоминающий звуки флейты, намного оживляет суровые высокогорные пейзажи.
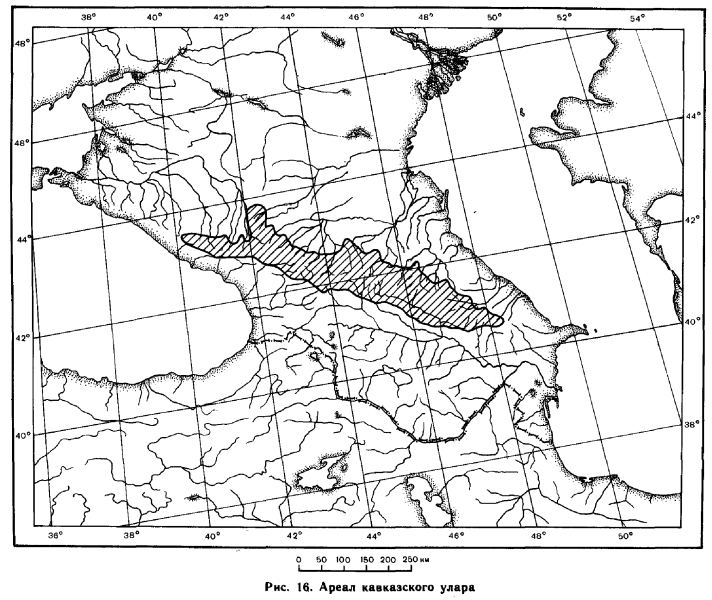
К северу от Главного хребта он найден на горах Бермамыт и Ханжал, южнее Кисловодска (кол. ЗИН АН СССР) и даже в верховьях р. Подкумок, где максимальная высота Бокового хребта составляет всего 2317 м над ур.м. (Левицкий, 1956). Самые западные точки — горы Оштен и Фишт (2808 и 2852 м над ур.м.), где они наблюдались, но добыты не были (Левицкий, 1956).
Видимо, улары доходят до этих вершин нерегулярно, поскольку есть указания на отсутствие этих птиц на названных вершинах (Аверин, Насимович, 1938). Восточной границей являются восточные склоны массива Базардюрю—Шахдаг—Бабадаг (Базиев, 1978). По южным склонам кавказский улар также опускается до указанных выше высот, но не ниже, и именно по этой причине не проникает на сравнительно низкие гребни Сурамского хребта, соединяющего Большой и Малый Кавказ. В целом же южная граница проходит севернее пос. Красная Поляна, захватывает верховья рек Мзымты и Бзыби, северную половину Кодорского хребта, Сванетский и Лечхумский хребты, верховья бассейна Риони, проходит севернее стыка Рачинского и Сурамского хребтов (встречен в июне в достаточном числе на высоте около 3000 м в 6 км севернее Квайсы, — по устному сообщению О. Р. Потаповой), захватывает верховья левых притоков Куры и верховья Арагви и тянется севернее Кахетинского и Картлийского хребтов на восток, вдоль южного склона Главного хребта, до вершины Бабадаг. В очерченных границах имеется ряд изолированных популяций, населяющих отдельные поднятия Скалистого хребта (Базиев, 1978).
Численность. Средняя численность птиц в альпийском поясе колеблется от 0.5 до 19.1 на 100 га, тогда как плотность на территориях, наиболее удобных для обитания птиц, варьирует от 1.5 до 47.9 птиц на 100 га, т.е. эти птицы способны создавать довольно значительную плотность, но в основном только осенью и зимой. В период гнездования плотность населения в оптимальных местах составляет от 12 до 31.5 птиц на 100 га.
Такие изменения плотности населения уларов в разные сезоны объясняются большей рассредоточенностью птиц в весенний период, когда у уларов начинает проявляться территориальное поведение (Базиев, 1978).
Сказывается также и крайняя неравномерность распределения птиц в различных частях ареала. В среднем плотность населения весной составляет в благоприятных местах обитания 20.5 птиц на 100 га или же 8.2 птиц на 100 га альпийского пояса в целом, а осенью соответственно 34.9 и 13.9 птиц на 100 га. Общая же численность кавказского улара составляет, исходя из указанной плотности, 164 тысячи птиц весной и 278 тысяч осенью (Базиев, 1978). На Западном Кавказе численность уларов заметно ниже и составляет, например в Тебердинском заповеднике, весной 3.3—6.6, а осенью — 4.6—8.5 особей на 100 га (Ткаченко, 1966).
Многолетние колебания численности отмечены в Тебердинском заповеднике, где общее число птиц изменялось в осенние сезоны от 700 до 1330 (Ткаченко, 1966). Вполне возможно, что численность на каждой конкретной территории может зависеть от перемещений птиц, поскольку улары очень подвижны, но названный автор склонен считать причинами таких колебаний характер весны, т. е. погодные факторы. В поздние вёсны, следующие за многоснежными зимами, когда снег долго скрывает обычные места гнездования, птицы поздно приступают к размножению, а некоторые вообще не гнездятся.
Питание пуховых птенцов состоит в основном из нежных листьев и верхушечных побегов альпийских трав, к которым впоследствии добавляются все новые и новые, причем более грубые части тех же растений, которые используются и взрослыми. Здесь опять же на первое место выходят листья, соцветия и незрелые бобы остролодочника, листья, коробочки и бутоны колокольчика, листья и незрелые плоды астрагалов, колоски и листья мятликов — все они вместе составляют 56 % рациона птенцов и молодых птиц. И опять обращает на себя внимание полное отсутствие насекомых в питании как самых маленьких птенцов, так и молодежи.
Весеннее оживление начинается в марте, разбивка на пары — с последней декады этого месяца, что совпадает с началом вегетации альпийской растительности. Брачный ритуал самца сравнительно прост и сводится к постоянному сопровождению самки, занятой кормежкой, более активному и нередко сдвоенному свисту, звучащему в таких случаях как особая брачная песня (Базиев, 1978), и к ряду поз ухаживания. Самец временами начинает кружить перед остановившейся самкой, поднимая вверх раскрытый хвост, опуская до земли крылья и распушивая все оперение (Noska, 1896). Ткаченко (1966) наблюдал своеобразный токовой полет этих птиц: вскоре после захода солнца пара уларов перелетала с одной стороны ущелья на другую (примерно 300 м) и через несколько минут обратно, причем во время полета самец держался на 1—2 м позади самки и, скользя на неподвижных крыльях, несколько раз высоко и резко поднимал вверх голову, одновременно слегка опуская вниз концы крыльев.
Повторив такой перелт несколько раз, птицы спарились. Процесс разбивки на пары и распределения по гнездовым территориям сопровождается драками между самцами, гоняющимися друг за другом. Позы и приемы у дерущихся птиц такие же, как и у домашних петухов. В течение всего апреля самцы очень активны и голосовые реакции их в это время особенно разнообразны.
Поведение пары уларов во время откладки яиц весьма специфично. Пробуждаются птицы за 1 ч 20 мин до восхода солнца и почти сразу же летят вниз, на кормежку. В дальнейшем в течение всего дня самка интенсивно кормится, за исключением краткого отдыха в середине дня. Самец же очень активен — бегает вокруг самки, часто свистит и усиленно кормится только тогда, когда самка отлучается к гнезду для откладки очередного яйца.
Гонады самцов достигают максимального развития в середине апреля (масса левого семенника 6 г), и в это же время происходит спаривание, а в третьей декаде апреля самки начинают откладывать яйца. Как и у остальных видов уларов, гнездо устраивается, как правило, на склоне южной, юго-восточной или юго-западной экспозиции, обязательно под прикрытием скалы, камня или кочки, в скальной нише или на скальном карнизе. В грунте для гнезда выкапывается неглубокая ямка, которая выстилается сухим растительным материалом, в основном листьями и стеблями разных трав, взятых в непосредственной близости от гнезда. Если поблизости растет много мха, то и он используется для выстилки.
В тех же случаях, когда вокруг нет никакой растительности (например, при гнездовании в скальной нише), скудная подстилка состоит исключительно из перьев насиживающей птицы (небольшое их количество встречается в любом гнезде). Внутренний диаметр лотка 17—24.5 см, глубина — 4—8 см.
Окраска яиц зеленовато-голубая, с различным количеством коричневых пятнышек и крапинок, то равномерно разбросанных по всей поверхности, то сконцентрированных на тумж конце. Размеры яиц заметно варьируют.
Средний размер (п = 42) 67.12 х 45.71 при максимальном размере 68.9 х 47.8 и минимальном 63.5 х 42.2. Средняя масса свежего яйца 77.62 (60.4—84.2). Число яиц в кладке 5—8, в среднем 6 (по 7 кладкам). Для того, чтобы отложить полную кладку, самке требуется от 9 до 15 дней, поскольку каждое яйцо откладывается через день, а иногда и через два (Базиев, 1978). Режим насиживания довольно напряженный, самка отлучается на кормежки в среднем дважды в сутки, утром и вечером, примерно на 20 мин, и в некоторые дни может даже не кормиться, оставаясь все время в гнезде. В ясную погоду под прямыми лучами солнца самка явно страдает от жары: она приподнимает перья и часто дышит широко открытым ртом. Будучи неоднократно потревоженными, птицы гнезд все же не бросали (Ткаченко, 1966). Незадолго до вылупления птенцов самка может подпускать человека вплотную и даже выдерживает прикосновение рукой. Длительность насиживания в 2 прослеженных случаях составила 28 дней. Вылупление птенцов происходит довольно дружно, в течение нескольких часов. Масса однодневных птенцов 58.3—65.4, причем масса остаточного желтка у птенца через 7 ч после вылупления составляла 11.5, а у птенца в возрасте 30 ч, еще не приступавшего к самостоятельному питанию, уже было некоторое количество подкожного и внутреннего жира, а также оставался еще нерассосавшийся желток. При вылуплении в неблагоприятных погодных условиях (холод, снегопады) выводок может оставаться в гнезде, обогреваемый самкой, более 2 сут и сохраняет при этом хорошую жизнеспособность.
Рост и развитие птенцов происходят очень интенсивно. В возрасте 7 дней они уже начинают перепархивать. Молодые птицы достигают размеров взрослых птиц к концу 3-го месяца жизни, а в 4-месячном возрасте почти сравниваются с ними по массе. Половой диморфизм в массе и размерах начинает проявляться примерно с месячного возраста. Так, масса самок составляет на 15-й день жизни 175, на 30-й — 360, на 45-й— 675, на 60-й —760, на 75-й— 1340, на 90—100-й— 1470 и на 1-25-й — 1550 (масса взрослых самок в это время 1575). У самцов на 30-й день масса составляет 390, на 60-й — 930, на 75-й — 1620, на 90—100-й — 1900 и на 125—130-й — 1600, при массе взрослых самцов в это время 1840 (Базиев, 1978). В Тебердинском заповеднике масса молодых самок 1 октября составляла 1340 при средней массе взрослых самок 1693. Максимальное прибавление в массе молодых птиц наблюдалось здесь в начале августа и составляло 18 г в сутки (Ткаченко, 1966).
С первых же дней жизни птенцы при тревоге разбегаются и затаиваются в камнях. Тревожное квохтание самки звучит, как «кок-кок-кок-кок-кок. . .», а при созыве выводка или призывая отбившегося птенца она издает своеобразную позывку «чуиии-чу-чу». Молодые самцы в возрасте 90 дней уже способны издавать все звуки, присущие взрослым самцам.
Повторных кладок взамен утерянных у кавказского улара, видимо, не бывает. Во всяком случае 2 самки, у которых были взяты кладки на 2-й день насиживания, к повторной кладке не приступили.
Поведение самцов в период насиживания очень напоминает таковое тундряной куропатки. В первые дни они находятся вблизи своих самок, занятых насиживанием и обычно сопровождают их на кормежках, возвращаясь вместе с ними к гнезду и нередко усаживаясь на скалу почти рядом, в 10—12 м. Там самцы могут свистеть и вести себя достаточно возбужденно, как и в период спаривания. Однако в это время никем не наблюдались ни защита самцом гнездового участка, ни другие формы территориального поведения. За 10 дней до появления птенцов самцы покидают гнездовые участки, объединяются в небольшие группы по 3—7 птиц и ведут кочевой образ жизни, забираясь на время линьки в самые высокие и труднодоступные места. По-видимому, они объединяются там с холостыми самками и самцами — первогодками, поскольку в таких местах можно встретить стаи по 25 птиц (Беме, 1926; Насимович, 1947).
Голосовые реакции те же самые, хотя указывается на то, что свист его более чист и звонок, чем у остальных уларов и слышен почти за километр (Сушкин, 1914). Нам же, например, никаких отличий в свисте кавказского и гималайского уларов подметить не удалось. В пределах ареала кавказский улар не может быть спутан ни с одной другой птицей, а вместе с другими видами уларов он нигде не встречается.
Величина суточного рациона определена Базиевым приблизительно в 400—450 г сырой массы (сухие стебли, колоски, семена альпийских трав). Приблизительную калорийность его определить трудно, поскольку неизвестно процентное соотношение отдельных компонентов корма, в том числе такого высококалорийного, как семена. Средняя масса содержимого зоба в зимний период составляет 62.6 г (Базиев, 1978). Птицы держатся всю зиму небольшими стайками, состоящими из 5—15 птиц обоего пола, как молодых, так и старых. Изредка встречаются более крупные стаи, до 30 птиц.
Сезонные перемещения. Обитая постоянно в течение всей своей жизни в одних и тех же горных группах, улары постоянно совершают незначительные вертикальные перемещения в поисках оптимальных условий обитания в тот или иной сезон года. Ранней весной, когда начинают освобождаться от снега самые нижние части субальпийского пояса вдоль верхней границы леса, улары спускаются сюда, привлекаемые травянистой растительностью, начавшей вегетацию. Поэтому в марте кавказские улары спускаются наиболее низко, до высот 2300—2400 м над ур. м. По мере продвижения вверх весны, а вместе с ней и пояса вегетации, начинают подниматься выше и улары. В середине лета они обитают в самых верхних частях альпийского пояса, близ вечных снегов, на высотах 3500—4000 м над ур.м. Осенью после первых снегопадов начинается обратное движение вниз, но задерживается на тех же высотах, где птицы гнездились — в средних и нижних частях альпийского пояса; здесь улары остаются на всю зиму. Масштабы вертикальных перемещений уларов в различных районах разные: они зависят от высоты горных гребней, от наличия или отсутствия вечных снегов и от положения снеговой линии. На Западном Кавказе вертикальные кочевки происходят в пределах высот 2200—3000 м над ур.м., в центральной части Кавказа — 2400—3800, а в восточной — 2400—4000 м над ур.м. (Базиев, 1978).
Рост самого дистального 10-го первостепенного махового оканчивается в возрасте 110 дней, а последнее взрослое первостепенное маховое, сменяющее ювенальное — 7-е, завершает свой рост на 140-й день жизни птенца.
Второстепенные маховые сменяются от 3-го внутрь, 2-е сменяется одновременно с 10-м, а первое — с 15-м. Последним, в возрасте 150 дней, заканчивается рост самого внутреннего, 16-го, второстепенного махового. Рулевые перья сменяются, начиная от 7-й пары (считая от центральной), сначала внутрь (6-я и 5-я пары), а затем в возрасте 40 дней и кнаружи. Последней, в возрасте 80 дней, сменяется центральная пара, но позже всех заканчивает свой рост наружная 10-я пара (к 110-му дню жизни птенца).
Контурное оперение ювенального наряда начинает сменяться на дефинитивное в возрасте 40 дней; сначала на верхней части груди, спине и крыльях, затем уже на боках, пояснице, брюхе, бедрах и в последнюю очередь на шее и голове. Тем не менее, оперение на голове вырастает быстрее, чем на брюхе и бедрах.
Следующая линька, которая затем повторяется ежегодно, — полная годовая. У самцов она начинается на 40 дней раньше, чем у самок, с выпаданием 1-го первостепенного махового, что происходит 15—20 мая.
Вслед за этим все первостепенные маховые сменяются в строгой последовательности вплоть до 10-го, которое заканчивает свой рост в первых числах ноября. За этот период, охватывающий 170 дней, происходит полная смена всего остального оперения. Первые маховые кисти растут быстро; выпадая с интервалами в 5—6 дней, они вырастают за 27—28 дней, но чем ближе к дистальным маховым, тем скорость смены и роста перьев медленнее. Так, 10-е маховое выпадает только через 18 дней после выпадения 9-го, а рост его продолжается почти 70 дней. Смена второстепенных маховых происходит более сложно, обычно из 2 центров. Сначала почти одновременно с 6-м первостепенным маховым выпадает 14-е и несколько позже 3-е. Затем в обе стороны от 14-го сменяются 15-е и 16-е, 13-е и 12-е.
Одновременно от 3-го второстепенного махового перья сменяются внутрь вплоть до 11-го, и только на последних этапах одновременно с 9-м сменяется 2-е и с 10-м — 1-е второстепенные маховые. Эта общая картина смены второстепенных маховых имеет довольно частые индивидуальные отклонения. Например, 2-е может выпадать раньше 1-го или одновременно с ним.
Рулевые перья начинают сменяться одновременно с 6-м первостепенным маховым, причем смена правых и левых рулевых происходит асимметрично. Только 9-я и 10-я пары выпадают и сменяются одновременно.
Но в целом линька хвоста идет так же, как и у молодых птиц, из 2 центров, начиная с перьев 5-й пары. Отрастание центральной пары заканчивается к 25 сентября, через 130 дней после начала смены первостепенных маховых, а крайней пары — к 15 октября, т. е. через 150 дней. Контурное оперение у самцов начинает сменяться одновременно со сменой второстепенных маховых — сначала на боках тела, затем на спине и нижней части шеи, потом на зобу, груди, подхвостье и брюхе. Еще позже линяют перья на бедрах и голени и в последнюю очередь — на шее и голове. На голове последовательность смены перьев такова: кроющие уха, на затылке, темени, лбу, на щеках с уздечкой и на горле. Однако среди контурных перьев последними заканчивают свой рост перья на боках тела и бедрах.
Самки начинают линьку примерно на 45 дней позже самцов, спустя 3 недели после вылупления птенцов. Порядок линьки перьев крыла, хвоста и контурного оперения такой же, но линька протекает в гораздо более сжатые сроки и завершается практически одновременно с линькой самцов.
Отмечены случаи, когда у самок, сильно запоздавших с откладкой яиц, линька начиналась уже в процессе насиживания. Общая продолжительность смены маховых перьев у самок, а следовательно и всей линьки — 4 мес.
Подробнее о сокращениях можно прочесть здесь.
Песня
…
Позывка
…
Электроманок
…
Литература
• Коблик Е.А. 2025. Фауна птиц стран Северной Евразии в границах бывшего СССР: Списки видов.
• IOC World Bird List v15.1 (Updated February 20, 2025) by Frank Gill, David Donsker & Pamela Rasmussen.
• Н.Н. Балацкий. Птицы Северной Евразии. Таксономический список: август 2023 г.
• «Птицы СССР. Курообразные. Журавлеобразные». т.2. Р.Л. Беме, Н.П. Грачев, Ю.А. Исаков, А.И. Кошелев, E.Н. Курочкин, Р.Л. Потапов, А.К. Рустамов, В.Е. Флинт. Ленинград. Издательство «Наука». 1987 г. стр. 100-108.
Сайты
• zmmu.msu.ru
• www.balatsky.ru
• www.worldbirdnames.org
• Avibase by Denis Lepage
• Zoonomen by Alan Peterson
• Taxonomy by John Boyd
• сайт Василия Вишневского
• www.xeno-canto.org
• www.featherbase.info, www.pbase.com, www.macaulaylibrary.org