

Статус. Таёжная овсянка Ocyris tristrami (Swinhoe, 1870) (рис. 1) – обычный гнездящийся перелётный вид Приморья.
Распространение и численность. В подходящих местообитаниях эти овсянки гнездятся по всей материковой части Приморского края.
Для залива Петра Великого размножение одной пары отметили в ХХ веке в густом мелколесье острова Матвеева (Лабзюк и др. 1971), но позднее гнездящихся птиц на островах не находили (Назаров, Шибаев 1984; Назаров 2001; Тиунов 2004). В летний период овсянок встречали на острове Русский (Назаров 2004), но гнездование здесь не доказано.
Относительная численность этого вида в 1962-1971 годах в чернопихтово-широколиственных лесах заповедника «Кедровая падь» варьировала от 1.4 до 11.2 пар/км2 (Назаренко 1984). В 1997 и 2001 годах в долинных многопородных лесах среднего и нижнего течения рек плотность населения таёжных овсянок достигала 18 пар/км2 (Gamova 2002).
В 2008 году обилие птиц в заповеднике составляло: 1.3-5.1 пар/км2 – в широколиственных лесах; 4.1-38.5 – в хвойно-широколиственных лесах; 40.1-42.0 – в долине реки Кедровая; 1.9-36.0 – в поймах притоков; 0-1.8 – в долине реки Барабашевка; 0-1.7 пар/км2 – в разных вариантах пирогенного древесно-кустарниково-лугового комплекса (Курдюков 2014)

В лесах Борисовского плато в среднем течении реки Грязная таёжная овсянка – обычный гнездящийся вид: в середине мая 2019 года плотность населения достигла 16.2 ос./км2. В долинных местообитаниях, густо заросших рябинником рябинолистным Sorbaria sorbifolia, в марте того же года обнаружили несколько типичных для этого вида прошлогодних гнёзд (выстилка лотка включала только чёрные придаточные корни папоротника). В начале июля осмотрели 18 гнёзд, 5 из которых были пустыми (разорены, либо птенцы уже вылетели), 5 – свежевыстроенными, 5 содержали яйца разной степени насиженности, а в 3 гнёздах находились птенцы (Беляев и др. 2019). В долине реки Лиственничная в начале июня 2023 года этих овсянок мы не обнаружили. Вероятнее всего, их отсутствие здесь – результат крайне высокой плотности населения пятнистых оленей Cervus nippon – поскольку подрост и подлесок были полностью уничтожены этими копытными, овсянки не имели в долине Лиственничной гнездовых стаций. В бассейне реки Абрикосовка плотность населения таёжных овсянок также была довольно низкой и во второй половине мая 2024 она находилась в пределах от 2.3 до 8.9 ос./км2 (наши данные), что тоже может быть связано с угнетением подроста и подлеска в результате пастьбы пятнистых оленей.
В 1962-1971 годах в хвойно-широколиственных лесах Уссурийского заповедника плотность населения таёжных овсянок составляла 1.4-11.2 пары на 1 км2 (Назаренко 1984). По другим сведениям, в этом типе местообитаний данный показатель варьировал от 1.1 до 3.3 пар/км2 в 1962-1976 годах; от 10.7 до 33.9 – в 1998–2005; от 35.4 до 63.8 пар/км2 – в 2013-2016 годах (Курдюков 2017), то есть явно прослеживается тренд значительного роста численности. В долинных широколиственных лесах заповедника в мае-июле 2024 года плотность гнездования таёжных овсянок колебалась от 10.0 до 18.2 ос./км2 (наши данные).
В кедрово-широколиственных лесах лесного участка Приморского ГАТУ (=Приморская ГСХА) в окрестностях сёл Каймановка и Каменушка Уссурийского городского округа в мае-июне 2020 года относительная численность составляла 18.4-20.8 ос./км2, в 2021 – 10.0-15.3, в 2022 – 13.8, в 2023 – 16.9 ос./км2. В широколиственных лесах долины реки Комаровка в 2020 году плотность населения вида достигала 21.3 ос./км2, в 2022 – 7.2, в 2023 – 16.6 ос./км2. Во вторичных дубняках в северной части лесного участка в окрестностях села Раковка относительная численность была гораздо ниже: в 2020 году – 2.4 ос./км2, в 2022 – 0.5, в 2023 – 8.1 (наши данные).
В конце 1970-х годов в западных отрогах Сихотэ-Алиня (хребет Синий) в елово-кедрово-широколиственных, в кедрово-широколиственных и в широколиственных лесах относительная численность этих овсянок составляла 9.1-25.8; 18.1-32.2 и 15.6-22.2 ос./км2, соответственно (Кушнарёв 1984). На этой же широте, но в условиях малооблесённой Ханкайско-Раздольненской равнины таёжные овсянки редки и распространены очень локально. Впервые в гнездовой период беспокоящуюся самку обнаружили в дубняках Гайворонской сопки 30 июня 2002, причём её поведение свидетельствовало о возможном гнездовании (Глущенко и др. 2006б). В дальнейшем 23-26 мая 2013 здесь зарегистрировали 5 территориальных самцов и нашли 2 гнезда (Глущенко и др. 2016). Следует отметить, что в некоторых районах Южного Приморья в XXI столетии для этих овсянок выявили значительное смещение биотопических преференций в плане выбора мест размножения, в частности, нарастание численности в галерейных и дубовых лесах (Курдюков 2010). Скорее всего, появление таёжных овсянок на гнездовании на Приханкайской низменности является следствием проявления этого же феномена.
В переходных от смешанных к темнохвойным лесах истоков реки Уссури и в пихтово-еловых лесах истоков реки Большая Уссурка в 1962-1971 плотность населения составляла 12.0-20.0 и 5.4-10.4 пар/км2, соответственно, а в зеленомошных пихтово-еловых лесах истоков реки Уссури этот вид не наблюдали (Назаренко 1984). В среднем течении Большой Уссурки, в национальном парке «Удэгейская легенда», таёжная овсянка – малочисленный гнездящийся вид. В июле 2020 года встречаемость этих птиц составляла 0.14 ос./км маршрута (Беляев 2022). В начале июня 2021 года эта овсянка входила в число доминирующих видов в долинных кедровниках в районе слияния рек Арму и Большая Уссурка, а плотность её населения была 25.0 ос./км2 (Беляев, Коваленко 2023). В дубовых лесах в районе КПП «Корейский прижим» таёжные овсянки встречались значительно реже – 1.3 ос./км2 (наши данные).
В Лазовском заповеднике в гнездовые периоды 1974-1975 годов средняя численность таёжных овсянок в кедрово-широколиственном лесу урочища Америка составляла 2.5 пар/км2 (Лаптев 1984). В дубняках долины реки Перекатная в 1978 году обилие этих овсянок не превышало 0.2 ос./км2, доля в населении – 0.8 %; в долинном кедрово-широколиственном лесу в 1988 году – 5.0 пар/км2, в 1992 – 7.1 пар/км2, доля в населении 2.5 %. В долинном многопородном лесу в 1993 году плотность гнездования составляла 7.7 пар/км2, доля в населении – 2.7 %, в 1994 – 3.8 и 0.9 %, соответственно. В 2001 году в долинном лесу реки Перекатная относительная численность таёжных овсянок достигала 37.50±0.49 ос./км2 (Шохрин 2017).
В гнездовой период 2017 года в Сихотэ-Алинском заповеднике в дубняках, смешанных и хвойных лесах плотность населения этих овсянок составляла 46.0, 47.5-70.4 и 46.2 ос./км2, соответственно (Начаркин и др. 2018).
Местообитания. Среди гнездящихся овсянковых Приморья таёжная овсянка является единственным по-настоящему таёжно-лесным видом как по общему рисунку распространения, так и по глубине проникновения её поселений вглубь сомкнутых массивов смешанной и темнохвойной тайги (Михайлов 2014). По определению А.А.Назаренко (1984), таёжные овсянки – характерные обитатели хвойно-широколиственных и частично пихтово-еловых лесов южного Сихотэ-Алиня. В поясе горных ельников они поднимаются до высоты 1000 м над уровнем моря, причём здесь птицы локализованы в долинах горных ключей, так как экологически они связаны с нижним кустарниковым ярусом леса. Только на хребте Пидан эти овсянки достигают верхней границы ельников и заходят в каменноберёзовое криволесье (Назаренко 1971б).
На крайнем юго-западе Приморья, у вершины горы Высотная, распространены до 900 м над уровнем моря (Панов 1973), а на Борисовском (Шуфанском) плато верхний предел распространения составляет 700 м (Назаренко 2014). В Северном Приморье на широте реки Бикин эти овсянки отсутствуют в области лесостепи и мозаичного ландшафта с лиственными островными лесами приуссурийской полосы, включая сопочные дубняки и кедровые островные леса на марях, и заселяют только островные облесённые хребты со смешанными и хвойными лесами (например, хребет Стрельникова). Птицы обычны в долинных, приречных лесах и тайге гористых водоразделов вплоть до подгольцовых редколесий в области среднего и верхнего Бикина (Михайлов и др. 1998; Пукинский 2003; Михайлов, Коблик 2013).

В заповеднике «Кедровая падь» таёжная овсянка «населяет хорошо облесённые долины горных речек (практически от уровня моря) с широколиственными и хвойно-широколиственными лесами. Поднимается высоко на склоны, но также главным образом по ключам. Однако известны случаи гнездования в липово-широколиственных лесах на совершенно “сухих” местах. В дубовых лесах отсутствует» (Назаренко 1971а, с, 44). По нашим данным, в этом заповеднике, в долине реки Кедровой, это обычный гнездящийся вид, занимающий практически все типы леса с хорошо выраженным кустарниковым ярусом за исключением пихтовои кедрово-широколиственных парковых лесов верхнего течения. В среднем и нижнем течениях Кедровой эти овсянки преобладают в светлых смешанных лесах из ясеня маньчжурского Fraxinus manshurica, ильма долинного Ulmus japonica, ольхи пушистой Alnus hirsuta с хорошо развитым травяным ярусом (осоки Carex spp., сердечник белоцветковый Cardamine leucantha, калужница лесная Caltha silvestris). Степень участия хвойных пород (Pinus koraiensis и Abies holophilla) в древостое этих лесов невелика и составляет не более 5 %. Здесь птицы придерживаются узких участков вдоль рек и ключей с зарослями рябинника рябинолистного Sorbaria sorbifolia. Встречаются также на отмелях и галечниковых косах, поросших чозенией Chosenia arbutifolia, ивами Salix spp., тополем Максимовича Populus maximowiczii, рябинником рябинолистным и густым травянистым покровом из Urtica angustifolia, Carex sordida, Filipendula palmata (Gamova 2002).
В Лазовском заповеднике эти овсянки гнездятся «по разреженным участкам широколиственных ильмово-липовых многоярусных лесов с подлеском из жимолостей, барбариса, бересклетов» (Винтер, Мысленков 2011, с. 308).
В бассейне реки Бикин – это обычная птица, которая «предпочитает кедрово-лиственные леса с зарослями черёмухи и чубушника в нижнем ярусе, папоротниками и относительно бедным и редким разнотравьем.
Однако гнездится и в смешанных лесах других типов, а также в старых лиственных лесах. В верховьях реки поселяется на склонах сопок, сухих лиственничных марях на плато с островами из лиственницы, ольхи и берёз» (Пукинский 2003, с. 226).
В целом для размножения таёжные овсянки обычно избирают участки с наиболее плотными куртинами подлеска, где часто довольно близко друг к другу гнездятся 3-4 пары (Курдюков 2003). Обязательным условием для гнездования этого вида в Приморье является наличие разнотравно-кустарниковых зарослей, произрастающих вдоль горных рек и ключей (Кулешова 1968; Панов 1973; Назаренко 1979).
Некоторые варианты типичных гнездовых биотопов таёжных овсянок иллюстрируют рисунки 2 и 3.
В период сезонных перемещений таёжные овсянки встречаются повсеместно в лесах, в зарослях кустарников и разнотравья на равнинах и в горах. Численность мигрирующих птиц широко варьирует по годам.

Весенний пролёт. Весной в южной половине Приморского края эти овсянки обычно появляются со второй декады апреля или с начала мая (Белопольский 1950; Омелько 1956; Назаров 1964; Панов 1973; и др.) (табл. 1; рис. 4). Указание на встречу нескольких самцов в стайке желтогорлых овсянок Cristemberiza elegans на окраине Владивостока 31 марта 1977 (Назаров 2004) нам представляется ошибочным (за таёжных овсянок, на наш взгляд, были приняты овсянки-ремезы Ocyris rusticus, типичные для этого времени года и часто образующие в этот фенологический период общие стайки с пролётными желтогорлыми овсянками).

Промежуток между первым появлением самцов и самок составляет 8 дней (Панов 1973). Пролёт длится до конца мая, когда, по нашим данным, встречаются преимущественно первогодки. В бассейне реки Бикин таёжные овсянки в разные годы весной появлялись с конца апреля по середину мая (Пукинский 2003; Коблик, Михайлов 2013). Любопытно отметить, что в отличие от большинства других мигрантов, для этих овсянок здесь зарегистрировали тренд смещения сроков пролёта на более позднее время (Коблик, Михайлов 2013).
На островах залива Петра Великого основной пролёт таёжных овсянок проходит в первой половине мая (Лабзюк и др. 1971). В Южном Приморье заметное увеличение численности самцов отмечали в течение первой декады мая, а к её концу начинают встречаться самки, хотя пролёт продолжался и в третьей декаде месяца, когда наблюдали группы, состоящие из 3-4 птиц (Панов 1973).
В первой половине ХХ столетия в Лазовском заповеднике массовый пролёт наблюдали уже в конце апреля, когда встречали стаи численностью 70-100 особей (Белопольский 1950), но позднее таких крупных транзитных групп здесь не наблюдали, а основная миграция проходит в первой половине мая (Шохрин 2017; наши данные).

Негативный тренд численности пролётных таёжных овсянок отмечен на островах залива Петра Великого: если во второй половине ХХ столетия для Дальневосточного морского заповедника эту овсянку указывали как многочисленный пролётный вид (Назаров, Шибаев 1984), то в текущем столетии она оказалась здесь малочисленной (Глущенко, Коробов 2013, 2014; наши данные).
Миграция продолжается до конца мая. В окрестностях Уссурийска наиболее поздние встречи явно пролётных птиц датированы 25 мая 2003 и 2004 (Глущенко и др. 2019), а на Приханкайской низменности – 25 мая 1978, 1995 и 2006; 26 мая 1986; 27 мая 2007; 29 мая 1980; 30 мая 1972; 7 июня 2011 (Глущенко и др. 2006б; наши данные).
Гнездование. По нашим данным, песни самцов слышны с момента их появления весной до середины июня (рис. 5), но полностью они замолкают лишь в конце июля. По другим данным, самцы поют с начала второй декады мая по середину июля (Панов 1973).
Для бассейна Бикина Ю.Б. Пукинский (2003) указывает, что таёжные овсянки активно поют и во время интенсивного пролёта (конец апреля – первые числа мая), когда встречаются стайки из 3-10 особей, а в крону одного дерева слетается до 30-50 птиц, при этом они «одновременно торопливо поют вразнобой укороченные песни, которые издали звучат как непрерывный громкий щебет» (Пукинский 2003, с. 226). По нашим наблюдениям, на юге Приморья такое пение более характерно для овсянок-ремезов, тем более в апреле, когда таёжные овсянки даже в южных районах края встречаются достаточно редко.
По нашим данным, на юге Приморья до начала образования пар таёжные овсянки кормятся вместе по 4-6 особей, предпочитая участки, подвергшиеся палам. Самцы начинают устанавливать границы индивидуальных участков с 12-14 мая с помощью рекламирующего пения в нескольких точках территории и шумных погонь, в которых участвуют 2-3 самца. К этому времени некоторые пары, образованные из рано прилетевших птиц, уже приступают к спариванию и строительству гнёзд.
Первый пик песенной активности вида, сопряжённый с появлением самок, отмечен во второй декаде мая. Спаривание происходит в 5-10 м от гнезда и у некоторых пар может продолжаться до середины срока насиживания кладки (Gamova 2002). В среднем течении Бикина демонстративное пение начинается в первых числах мая, сразу после появления самцов на гнездовых участках, а в верховьях реки – с середины мая. Птицы регулярно поют до середины июня, а в конце этого месяца имеет место второй подъём интенсивности вокализации (Пукинский 2003).
Как показали наши наблюдения за мечеными птицами, для таёжных овсянок характерна компактность гнездовой территории, слабая дифференциация кормовых участков (один кормовой участок могут использовать несколько пар), стабильное число и строгая локализация песенных постов. Площадь охраняемой гнездовой территории колеблется от 1800 до 7200 м2, достигая наибольшей величины в смешанных многопородных долинных лесах.

Таёжные овсянки поют на 3-4 песенных постах, расположенных на деревьях, один из которых находится в нескольких метрах от гнезда, а остальные – в 30-60 м от него. В период строительства гнезда самец поёт тихие песни с большими межпесенными интервалами; нередко пение сочетается с кормодобывающей активностью. Самец чередует два варианта песен, различающихся заключительной частью. Перед спариванием в течение 5-10 мин он поёт серию коротких песен, а после него в течение 5 мин исполняет серию более длинных. После завершения постройки гнезда места наибольшей активности пары сосредотачиваются на песенных постах самца и на кормовом участке, в 100-120 м от гнезда.
В этот период пение активизируется и становится более громким, при этом повышается уровень тревожного поведения у гнездовой постройки.
В присутствии человека или в ответ на голос вороны самец не допевает концовку песни, а меняет один её вариант на другой, сменив песенный пост. В распоряжении самца находятся 3-4 варианта песен, которые он последовательно чередует в одной серии. Перед началом насиживания песенная активность достигает 90-150 песен в час, при этом непрерывное пение длится не больше 7 мин (Gamova 2002).
У таёжных овсянок имеется два репродуктивных цикла за сезон. Общая продолжительность гнездового периода конкретной пары колеблется от 30 до 67 дней в зависимости от наличия одной или двух кладок.
Овсянки из двух пар, находившихся под нашим наблюдением, второй раз загнездились через 1-1.5 недели после вылета птенцов из первых гнёзд. Вторые гнёзда птицы устраивали в 50 и 120 м от первых. Гнездовой период вида растянут примерно на два с половиной месяца: с середины мая по конец июля (табл. 2).
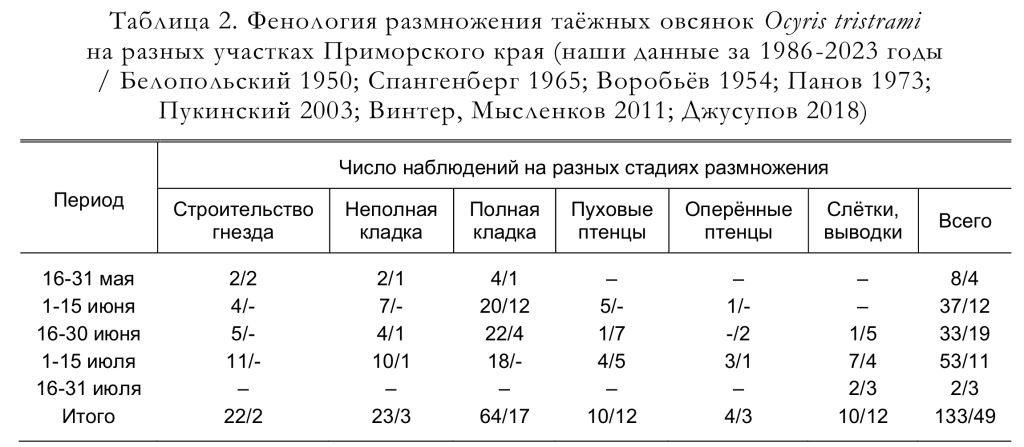
Самые ранние даты строительства гнёзд в 1961-1963 и 1980-1981 годах зафиксировали 26-27 мая (Панов 1973; Шибнев 1984). Гнездо строит самка, но материал приносят обе птицы, собирая его недалеко от постройки. Они прилетают к гнезду через каждые 2-5 мин, а материал самка укладывает в течение 1 мин (Gamova 2002).
В бассейне Бикина гнёзда располагались «среди травы, в розетках папоротников, на нижних ветках молодых ёлочек или подроста лиственных деревьев, на высоте 0.2-0.4 м. Характерны также постройки на невысоких кустах, преимущественно на спирее, рябинолистнике и элеутерококке, в 1-1.5 м от земли» (Пукинский 2003, с. 226). Данные других авторов и полученные нами сведения по расположению гнёзд таёжных овсянок оказываются сходными (табл. 3).

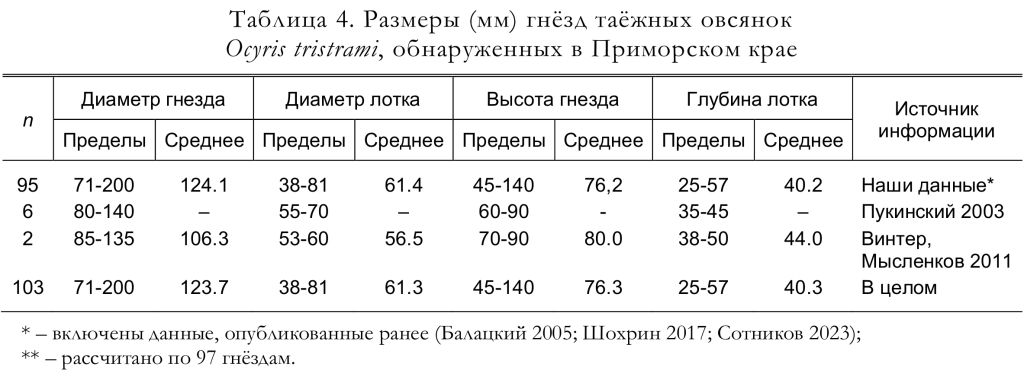
Типичные варианты размещения гнёзд иллюстрируют рисунки 6 и 7. Средняя высота расположения гнёзд в бассейне Бикина (n = 13) составила 0.8 м (Пукинский 2003). По нашим данным из разных районов края, высота расположения гнёзд (n = 117) составила 0-550, в среднем 87 см, при этом выше 150 см были устроены только 5 гнёзд.


Гнёзда, осмотренные Е.Н. Пановым на юге Приморья, были «построены однообразно, из сухой травы, очень характерна выстилка из чёрных стеблей папоротника Adiantum pedatum, не доходящая до края гнезда» (Панов 1973, с. 271-272). Два гнезда, найденные в Лазовском заповеднике в 1970-1971 годах, «выглядели рыхлыми и неаккуратными, их наружный слой состоял из сухих целых растений или стеблей злаков, неряшливо торчащих в разные стороны, средний – содержал сухие листья и стебли злаков, а также тонкие корешки древесных растений, лоток был выстлан чёрными “волосками” лишайника Usnea. В стенки первого гнезда были вплетены зелёные вайи папоротника, растущего под ним» (Винтер, Мысленков 2011, с. 308-309).
Наиболее подробное описание гнездовых построек таёжных овсянок приводит Ю.Б. Пукинский (2003, с. 226): «Поверх небольшого рыхлого основания из грубых стеблей злаков самка вьёт массивный внешний слой (до 2/3 гнезда по объёму) из того же материала. Особой толщиной выделяются его верхние края, где отдельные стебли достигают 40-55 см в длину. В ажурном и более аккуратно свитом среднем слое преобладают стебли злаков. Укладывая строительный материал, птица поворачивается то по, то против часовой стрелки; различная направленность укладки материала обеспечивает большую прочность стенок постройки. В них иногда вплетаются засохшие побеги кустарников, кедровая хвоя, прошлогодние листья папоротника. Просвечивающая выстилка лотка, толщиной у дна до 1 см, состоит из волосовидных чёрных корешков папоротников и грубой шерсти (преимущественно изюбря и лося)».
Размеры гнёзд приведены в таблице 4, вес внешней части постройки варьирует от 6.60 до 13.95 г, а масса выстилки – от 0.99 до 5.10 г (Gamova 2002).
По нашим данным, самки приступают к откладке яиц через 1-4 дня после завершения строительства гнезда. В бассейне реки Бикин, судя по находкам 12 гнёзд, завершение кладки приходится на первую декаду июня (8 случаев), реже (по 2) – на вторую или третью декады этого месяца (Пукинский 2003).
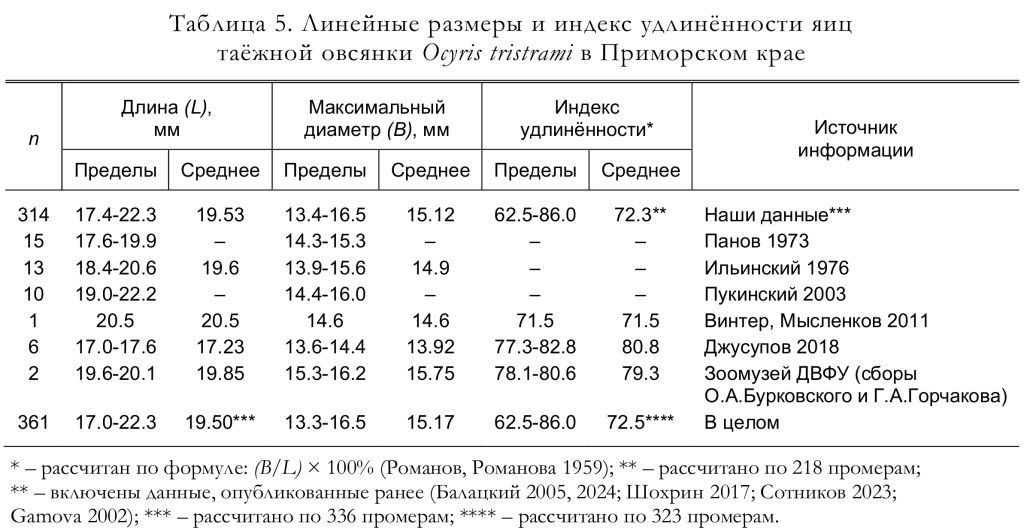
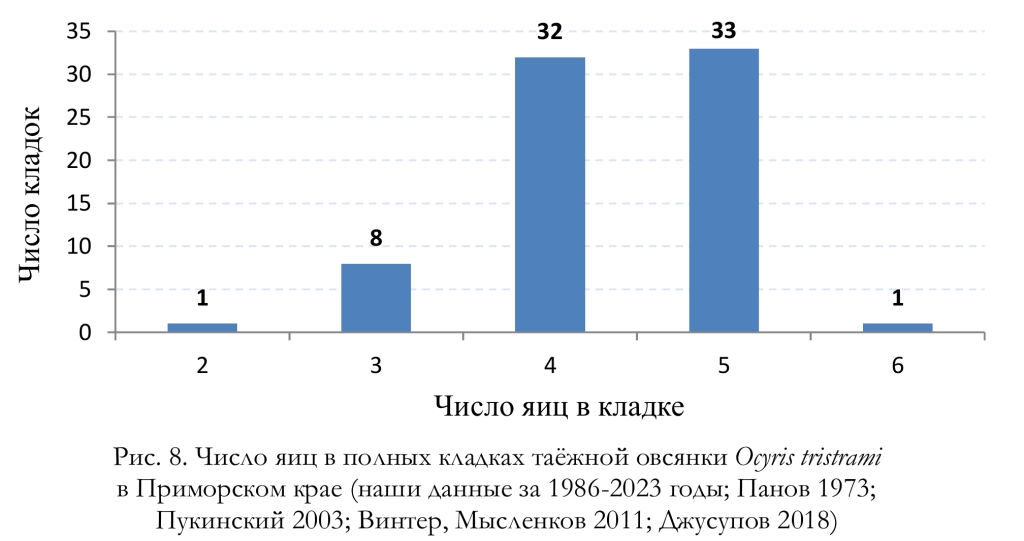
Все три кладки, найденные в Южном Приморье Е.Н. Пановым (1973), содержали по 5 яиц, так же, как и все 6 полных кладок, обнаруженных Ю.Б. Пукинским (2003) в бассейне Бикина. Согласно нашим материалам, число яиц в завершённых кладках варьировало от 2 до 5; чаще всего их было 4, а средняя величина кладки составила 4.22 яйца (n = 64). Следует отметить, что в кладках, осмотренных нами в мае и июне (n = 48), было 3-5 яиц, в среднем 4.42 яйца, а в июльских (вероятно, в большинстве случаев вторых) кладках (n = 16) было от 2 до 4 яиц, в среднем 3.63 яйца. Единственную известную для Приморского края кладку таёжной овсянки, состоящую из 6 яиц, коллектировал Е.П. Спангенберг в заповеднике «Кедровая падь» 31 мая 1968 (Джусупов 2018; Джусупов, Чупин 2022). Учитывая все приведённые выше данные, число яиц в полных кладках этого вида варьирует от 2 до 6 яиц (рис. 8), а средняя величина кладки составляет 4.37 яйца (n = 75). Линейные размеры, индекс удлинённости, вес и объём яиц приведены в таблицах 5 и 6.

По данным Е.Н. Панова (1973), основной фон яиц зеленоватый или зеленовато-палевый; глубокие фиолетовые, розовато-серые или розовато-коричневые пестрины густые и иногда они почти заслоняют основной фон на тупом полюсе или по всему яйцу, а поверхностные тёмно-бурые или чёрно-фиолетовые пестрины имеют вид палочек, запятых, зигзагов и нитей, слегка размытых по краям. Согласно другому описанию, «по грязно-зеленовато-белому фону скорлупы яиц двух кладок были изредка разбросаны чёрно-бурые точки и извилистые линии, относительно равномерные на яйцах одной кладки и сгущавшиеся на тупом полюсе – другой» (Винтер, Мысленков 2011, с. 309). По материалам Ю.Б. Пукинского (2003, с. 226), «скорлупа голубовато-белая, с рисунком из буровато-фиолетовых пятен, точек, сероватых штрихов и завитков; наиболее густо элементы рисунка расположены на тупом полюсе».
По нашим данным, окраска фона яиц однообразная, кремово-серая. Имеются глубинные серо-коричневые пятна, которые могут почти полностью заполнять поверхность яйца, и поверхностные тёмно-коричневые или чёрные точки, чёрточки и завитки. Глубинные отметины почти полностью покрывают поверхность (плотность заполнения – густая, сплошная), распределены равномерно или сгущаются на тупом конце, поверхностные – редкие (заполняют не более 5 % поверхности яйца) и сконцентрированы больше на тупом конце (Gamova 2002).
Для наглядности приводим коллажи, составленные из фотографий разных кладок таёжных овсянок из Приморского края, выполненных в условиях природы (рис. 9) и студии (рис. 10).
В насиживании кладки и обогреве птенцов участвуют оба партнёра (Панов 1973; Пукинский 2003; Винтер, Мысленков 2011; наши данные).
Ю.Б. Пукинский (2003) указывал, что кладку насиживает преимущественно самка, а самец регулярно сменяет её, проводя в гнезде до 25 % дневного времени. По нашим наблюдениям, время непрерывного насиживания у самки составляет 31-102, в среднем 66.8±28.1 мин, у самца – 22-54, в среднем 35.5±15.3 мин, а время отсутствия, соответственно – 22-54, в среднем 36.2±13.0 мин и 31=85, в среднем 66.3±30.6 мин.


Период насиживания в 16 гнёздах таёжных овсянок составил 11 сут (Gamova 2002), что соответствует наблюдениям Ю.Б. Пукинского (2003), по данным которого инкубация одной кладки продолжалась 11-12 сут.
Возле гнёзд с яйцами птицы ведут себя скрытно: улетают и прилетают в одном и том же направлении, быстро сменяют друг друга, не издают возле постройки позывок. Они перелетают у гнезда по дугообразной или прямой траектории низко над землёй. Насиживание очень плотное, и при появлении человека возле гнезда овсянки нередко оставляют кладку только тогда, когда наблюдатель касается рукой гнезда или самой птицы. После этого она спрыгивает с гнезда и перемещается поблизости или отлетает от него полукругом на 15-30 м. На относительно открытом месте овсянка начинает планировать параллельно движению человека. После этого садится на ветку дерева на высоте до 6 м и издаёт тревожные цыканье, которое при повышении уровня тревоги переходит в протяжный свист. Самка часто отводит от гнезда, перебегая по земле.
Отбежав на видное место, она сначала поднимает левое крыло, затем оба крыла, хвост широко разведён.
Несмотря на заметную роль в насиживании, во время отсутствия на гнезде самец активно участвует в защите территории от вторжений посторонних самцов. Нередко происходят драки с соседними самцами в 10-20 м от гнезда, повышается песенная активность. В период наибольшей суточной активности самец исполняет до 400 песен в час. Перед тем как поменяться с самкой на кладке он поёт рядом с гнездом до 6 песен, а слетая с него через минуту начинает петь, исполняя до 74 песен подряд в течение 4-10 мин. Лишь с середины срока насиживания активность пения снижается до 48-186 песен в час (Gamova 2002).
В Лазовском заповеднике вылупление птенцов в одном из гнёзд отметили 22 июня, а их вес в первый день жизни составлял 2.00, 2.60 и 2.65 г (Винтер, Мысленков 2011). В бассейне реки Бикин выход птенцов из яиц происходил во второй (5 случаев) и третьей (1) декадах июня и в первой декаде июля (1), а вес однодневных птенцов (n = 5) варьировал от 2.1 до 2.8 г (Пукинский 2003).
По нашим данным, в заповеднике «Кедровая падь» в 1997 и 2001 годах птенцы вылуплялись в период с 7 по 28 июня, а в другие годы это происходило 11 и 17 июня 1996 в бассейне среднего течения реки Бикин (Балацкий 2005), 8 июня 2017 в окрестностях посёлка Ясное (Чугуевский район), а также 2 и 6 июля 2017 в бассейне реки Грязная.
В заповеднике «Кедровая падь» в 1997 и 2001 годах отход яиц (n = 96) в 24 кладках составил 11.5 %. Общая успешность насиживания – доля вылупившихся птенцов (n = 85) от числа отложенных яиц (n = 96) – составила 88.5 % (Gamova 2002).
Однодневных птенцов описывали И.В. Ильинский (1976) и Ю.Б. Пукинский (2003). По нашим данным, птенцы (n = 67) имели опушение на 6 птерилиях: надглазничной (тёмно-серый, длина 6 мм), затылочной (серый, 5 мм), плечевой (серый, 7 мм), кистевой (серый, 5 мм), бедренной (светло-серый, 5 мм) и спинной (серый, 10 мм). У птенцов отсутствовал пух на локтевых птерилиях. Кожа оранжевая (в описании Пукинского она бледного розовато-жёлтого цвета). Яйцевой зуб серый, клювные валики белые (в статье Ильинского они жёлтые, а у Пукинского – бледно-жёлтые, в углах малиновые), клюв телесно-серый, его кончик тёмно-серый, ротовая полость бежевая (в описании Пукинского она интенсивнокрасная), ноздри овальные, параллельные коньку клюва. Размеры однодневных птенцов (n = 67), мм: клюв 4.0±0.8, плечо 8.5±1.2, кисть 9.0±0.4, бедро 11.5±1.5, голень 14.5±1.1, цевка 11.5±2.6 (Gamova 2002).
Интенсивность кормления птенцов, среднее время, затрачиваемое на поиски корма, а также доля участия в обогреве птенцов (для 4 гнёзд, общее время наблюдений 126 ч) у самки и самца примерно одинаковы.
За 1 ч овсянки прилетают с кормом 1-3 раза, принося птенцам по 1-4 пищевых объекта, и отлучаются в среднем на 32.8 мин. На обогрев птенцов они в среднем расходуют по 27.4 мин/ч. Отход птенцов (n = 85) в 26 гнёздах составил всего 3.5 %. В 3 гнёздах пропало по 1 птенцу в возрасте от 2 до 5 сут. Успешность размножения – доля вылетевших птенцов (n = 82) от числа отложенных яиц – составила 85.4%. Этот же показатель, рассчитанный по модифицированному методу Мэйфилда (Паевский 1985) составил 82 % (Gamova 2002).
У гнезда с птенцами самец ведёт себя более осторожно, чем самка.
Он незаметно покидает постройку: сначала вылетает вверх, а затем летит дальше по дугообразной траектории. При возвращении к гнезду не долетает несколько метров и подходит к нему по траве. В отличие от самца, самка залетает в гнездо сразу; покормив птенцов она может открыто перемещаться возле гнезда. По мере роста птенцов родители пролетают возле гнезда волнообразным и прямым полётом, низко над землёй. Первый тип полёта хорошо маскирует птицу, поскольку со стороны трудно определить первоначальное место взлёта и, следовательно, место нахождения гнезда. Прямой полёт выполняет транспортную функцию. Его основное преимущество – большая скорость, что особенно важно при доставке птенцам корма и очистке гнезда от экскрементов. Поведение самки при попытках человека взять птенца из гнезда может быть разным: она молча перелетает над рукой человека или нападает на неё.
Человека, удаляющегося от гнезда, самка преследует, зависая в воздухе на высоте до 80 см от земли. Самец чаще всего отбегает от гнезда и принимает позы угрозы. После вылупления птенцов самец почти не поёт, но часто издаёт одно-двухсложную позывку в 20-30 м от гнезда. Между кормлениями он поёт не более 38-58 песен в час. При тревоге самец оповещает самку об опасности тихой короткой серией песен с большими межпесенными интервалами (Gamova 2002).
Число птенцов в гнезде в 1 случае составило 3, в 1 – 4 и в 3 случаях – 5 (Пукинский 2003), то есть в среднем 4.4 птенца в выводке. По нашим данным, число гнездовых птенцов в разных выводках варьировало от 2 до 5, в среднем составляя 3.82 птенца (n = 11). Гнездовых птенцов таёжных овсянок (рис. 11) наблюдали с начала июня до середины июля, но судя по наличию сравнительно большого числа кладок в первой половине июля (табл. 2), гнездовые птенцы должны встречаться и во второй половине июля.

По наблюдениям Ю.Б. Пукинского (2003), в двух случаях птенцы находились в гнезде 8 и 9 сут. По нашим данным, они могут покидать гнездо уже на 8-е сут, но в случаях редкого появления человека у гнезда и снижения фактора беспокойства птенцы вылетают на 9-10-е сут. Продуктивность размножения довольно высокая: на одну успешно гнездившуюся пару (n = 22) приходится 3.7 слётка.
В бассейне Бикина птенцы оставили гнёзда 23, 26 июня и 17 июля, а большинство встреч плохо летающих слётков, которых кормили родители, приходилось на третью декаду июня (Пукинский 2003).
Только что оставивший гнездо слёток имеет буровато-серый клюв с жёлтыми клювными валиками, бледно-розовую ротовую полость, розовато-серую цевку. Размеры таких птиц (n = 70), мм: длина клюва 9.0±0.3, плеча 20.2±0.6, кисти 17.0±0.8, бедра 22.0±1.4, голени 29.2±1.5, цевки 22.0±1.3. Длина перьев (n = 40), мм: лобных 1.7±0.2 (чёрные с бледно-жёлтыми пестринами по краю пера), теменных 0.5±0.1 (грязно-белые), шейных 9.0±1.2 (чёрные со светло-коричневыми концами по краю), горловых 8.0±0.9 (бледно-жёлтые со светлым центром), брюшных 11.0±1.8 (бледно-жёлтые), первостепенных маховых 26.0±2.6 (тёмно-бурые с рыжим по внешней части опахала), спинных 13.0±3.1 (рыжие с чёрным по центру), копчиковых 5.5±0.6, голенных 3.2±0.7, бедренных 4.5±0.5 (светло-серые), брюшных 14.0±1.7 (грязно-жёлтые).
Слётков таёжных овсянок (рис. 12) наблюдали со второй половины июня до конца июля.

На четвёртый день слётки становятся более заметными и могут перелетать низко над землёй на небольшие расстояния, но предпочитают прятаться в траве. При тревоге они уже издают частую позывку, взлетают и садятся на веточки деревьев на высоте до 1 м. Возле недавно вылетевших птенцов в присутствии человека самка прыгает по земле, перемещаясь параллельно его движениям с распушённым брюхом и приподнятым оперением головы. Самец как правило находится на дереве в той же позе, что и самка, издавая тревожный свист. На девятый день слётки уже довольно хорошо летают. В этом возрасте они удаляются на 80 м от гнезда и находятся под присмотром самца. Возле недавно покинувших гнездо молодых овсянок он издаёт протяжный свист, который в присутствии человека становится более отрывистым. Активность пения в это время невелика (n = 13): с 12-13 ч – 78 песен/ч. Пение может прерываться смещёнными чистками оперения груди и плеч, при этом перья головы приподняты. Хотя песенные посты соседних самцов находятся на расстоянии не менее 100 м один от другого, при возникающих конфликтах самцы могут петь рядом один и тот же тип песни. На дальнем расстоянии друг от друга характерно антифональное пение разных типов песни. Поведение самца очень характерно: он взлетает с земли или присады вверх и поёт в воздухе.

Через две недели после оставления гнёзд птенцы становятся самостоятельными (Gamova 2002), что приходится на вторую половину июня и более поздние сроки (табл. 2; рис. 13). Так, в среднем течении реки Большая Уссурка неразбившиеся выводки мы наблюдали в 20-х числах июля 2020 года (Беляев 2022).

Осенние миграции. Послегнездовые кочёвки таёжных овсянок начинаются в августе, а заметный осенний пролёт проходит с первых чисел сентября, когда птицы попадаются в несвойственных им местообитаниях, и заканчивается в первой половине октября (Панов 1973). По результатам массового отлова птиц паутинными сетями, проводимого в окрестностях Лазовского заповедника в 2001-2013 годах, начало осенней миграции отметили 3 сентября 2005, а её окончание – 16 октября 2004 (Шохрин 2017). В период массового кольцевания птиц в Красноармейском районе (бассейн реки Большая Уссурка) в 2002 году осенняя миграция взрослых птиц отмечалась с 12 по 30 сентября, а молодых – с 7 сентября по 9 октября (данные С.Г. Сурмача и Т.В. Гамовой). Наиболее поздние встречи таёжных овсянок обычно происходят в разные числа октября (табл. 7; рис. 14).

В диссонанс с этими материалами вступают данные Л.О. Белопольского (1950), согласно которым пролёт таёжных овсянок в Лазовском (Судзухинском) заповеднике в 1940-х годах якобы проходил в первой и, отчасти, во второй декадах ноября, а также указан случай добычи самца 10 декабря 1944. Эти данные ошибочны, поскольку в Зоологическом музее Московского университета, где хранятся сборы Л.О.Белопольского, таёжных овсянок, добытых этим автором нет, зато есть 2 апрельских и 1 октябрьский экземпляры овсянок-ремезов, на этикетках которых указано «Emberiza tristrami» (письменное сообщение Я.А.Редькина).
Питание. Таёжные овсянки обычно собирают корм на земле и несколько реже – на кустарниках и в кронах деревьев. Среди содержимого 19 желудков добытых птиц преобладали гусеницы, хотя и растительные корма играли существенную роль в питании этих птиц даже в середине лета, составляя от 20 до 100 % от общего объёма пищевого комка (обнаружены в 12 желудках) (Назаров и др. 1979).
Наши наблюдения показали, что в летний период таёжные овсянки питаются преимущественно различными насекомыми и их личинками, реже поедают пауков (рис. 15.1) и моллюсков (рис. 15.2), а во время миграций в их питании присутствуют и растительные корма (рис. 15.3).
Неблагоприятные факторы, враги, гибель. Одну кладку из 5 яиц уничтожила большеклювая ворона Corvus macrorhynchos. Ещё одно гнездо с неполной кладкой птицы бросили из-за сильного ливня.
На многих участках национального парка «Земля леопарда» из-за высокой численности пятнистых оленей исчезает подлесок и формируются парковые леса, не пригодные для гнездования таёжных овсянок.


В окрестностях Лазовского заповедника таёжных овсянок отмечали в питании перепелятников Accipiter nisus и малых перепелятников A. gularis (Шохрин 2008, 2017). В Уссурийске одиночную особь нашли 20 сентября 2020 разбившейся об оконное стекло (Беляев и др. 2020).

Для Северного Приморья в качестве гнездового паразита таёжной овсянки известна обыкновенная кукушка Cuculus canorus (Пукинский 1978). Так, в среднем течении реки Бикин 18 июня 1974 в гнезде с незаконченной кладкой овсянки обнаружили яйцо этой кукушки, имеющее окраску, соответствующую окраске яиц хозяев, из которого вылупился и был благополучно выкормлен кукушонок (Пукинский 2003).
В этом же районе в окрестностях села Красный Яр 26 июня 2000 в гнезде желтогорлой овсянки обнаружили яйцо обыкновенной кукушки, окраска которого соответствовала окраске яиц таёжной овсянки (Глущенко и др. 2024) (рис. 16). Там же 2 июля 2000 в гнезде таёжной овсянки нашли яйцо глухой кукушки Cuculus optatus (рис. 17), которое явно попало в него по ошибке.
В 2017-2023 годах с отловленных в паутинные сети таёжных овсянок сняли мух-кровососок Hoppoboscidae трёх видов: Ornithoica momiyamai (41 особь), Ornithoica unicolor (1) и Ornithomya avicularia (12) (Nartshuk et al. 2023; наши данные).
Литература
• Балацкий Н.Н. 2005. К авифауне верхнего течения Бикина // Рус. орнитол. журн. 14 (278):
98-103. EDN: IJVUSN.
• Балацкий Н.Н. 2024. Гнёзда птиц Сибири и сопредельных регионов: справочник. Новосибирск, 4: 1-532.
• Белопольский Л.О. 1950. Птицы Судзухинского заповедника (воробьиные и ракшеобразные)
// Памяти академика П.П.Сушкина. М.; Л.: 360-406.
• Беляев Д.А. 2022. Предварительные данные о населении птиц бассейна реки Большая Уссурка (Национальный парк «Удэгейская легенда», Приморский край) // Вестн. ИрГСХА 3 (110): 45-63.
• Беляев Д.А., Глущенко Ю.Н., Горбуля А.А. 2020. Гибель птиц в Уссурийске (Приморский край) от столкновения с оконными стёклами // Амур. зоол. журн. 12, 1: 71-79.
• Беляев Д.А., Глущенко Ю.Н., Коробов Д.В., Тиунов И.М. 2019. Птицы бассейна верхнего течения р. Грязная (национальный парк «Земля леопарда») // Биота и среда заповедных территорий 4: 65-85.
• Беляев Д.А., Коваленко Д.Б. 2023. Данные о населении птиц бассейна реки Большая Уссурка (национальный парк «Удэгейская легенда») по результатам учётов 2021 года // Лесное хозяйство: материалы 87-й науч.-техн. конф. Минск: 49-53.
• Винтер С.В., Мысленков А.И. 2011. О птицах Лазовского заповедника // Сомовская библиотека. Вып. 1. Экология птиц: Виды, сообщества, взаимосвязи. Тр. науч. конф., посвящ. 150-летию со дня рождения Н.Н. Сомова (1861-1923). Харьков: 267-323.
• Воробьёв К.А. 1954. Птицы Уссурийского края. М.: 1-360.
• Глущенко Ю.Н., Коробов Д.В. 2013. Авифаунистические исследования на о-ве Фуругельма (Японское море) весной 2013 г. // Животный и растительный мир Дальнего Востока 2 (20): 9-16.
• Глущенко Ю.Н., Коробов Д.В. 2014. Авифаунистические исследования на крайнем юго-западе Приморского края весной 2014 г. // Животный и растительный мир Дальнего Востока 2 (22): 6-14.
• Глущенко Ю.Н., Коробов Д.В., Харченко В.А., Коробова И.Н., Глущенко В.П. 2019. Птицы – Aves // Природный комплекс Уссурийского городского округа; современное состояние. Владивосток: 151-301.
• Глущенко Ю.Н., Липатова Н.Н., Мартыненко А.Б. 2006а. Птицы города Уссурийска: фауна и динамика населения. Владивосток: 1-264.
• Глущенко Ю.Н., Нечаев В.А., Редькин Я.А. 2016. Птицы Приморского края: краткий фаунистический обзор. М.: 1-523.
• Глущенко Ю.Н., Шибнев Ю.Б., Волковская-Курдюкова Е.А. 2006б. Птицы // Позвоночные животные заповедника «Ханкайский» и Приханкайской низменности. Владивосток: 77-233.
• Джусупов Т.К. 2018. Оологические сборы Е.П. Спангенберга на юге Приморья, в центральной части, на севере и северо-востоке России // Selevinia 26: 107-129.
• Джусупов Т.К., Чупин И.И. 2022. Каталог оологической коллекции Института систематики и экологии животных СО РАН. Новосибирск: 1-170.
• Ильинский И.В. 1976. К биологии размножения таёжной овсянки (Emberiza tristrami) //
Вестн. Ленингр. ун-та 9: 42-48.
• Коблик Е.А., Михайлов К.Е. 2013. Изменения сроков прилёта птиц в бассейне реки Бикин
(север Приморского края) в 1990-е годы по сравнению с 1970-ми // Рус. орнитол. журн. 22 (948): 3341-3347. EDN: RNGVQB.
• Кулешова Л.В. 1968. Черногорлая овсянка (Emberiza tristrami Swinhoe) в Среднем Сихотэ-
Алине // Науч. докл. высш. школы. Биол. науки 12: 23-30.
• Курдюков А.Б. 2003. Мозаично-ярусная организация лесных ценозов как фактор биологического разнообразия птичьих сообществ юга Уссурийского края // Современные проблемы орнитологии Сибири и Центральной Азии. Улан-Удэ, 1: 103-107.
• Курдюков А.Б. 2010а. Глубокая динамика численности и смены предпочтений в использовании местообитаний у трёх модельных видов лесных птиц северо-восточной окраины Восточно-Маньчжурских гор // 9-я Дальневост. конф. по заповедному делу. Владивосток: 228-233.
• Курдюков А.Б. 2010б. Население птиц Верхнеуссурийского биогенотического стационара, пояса среднегорий Южного Сихотэ-Алиня // Рус. орнитол. журн. 19 (548): 191-221. EDN: KYKIYN.
• Курдюков А.Б. 2014. Гнездовые орнитокомплексы основных местообитаний заповедника «Кедровая Падь» и его окрестностей: характер размещения и состояние популяций, дополнения к фауне птиц (материалы исследований 2008 года) // Рус. орнитол. журн. 23 (1060): 3203-3270. EDN: SWMORL.
• Курдюков А.Б. 2017. Население птиц девственных неморальных хвойно-широколиственных лесов Южно-Уссурийского края: более полувека наблюдений // Динамка численности птиц в наземных ландшафтах. М.: 77-86.
• Кушнарёв Е.Л. 1984. Антропогенные сукцессии орнитосообществ и территориальные связи
местообитаний западного Сихотэ-Алиня // Фаунистика и биология птиц юга Дальнего Востока. Владивосток: 71-78.
• Лабзюк В.И., Назаров Ю.Н., Нечаев В.А. (1971) 2020. Птицы островов северо-западной части
залива Петра Великого // Рус. орнитол. журн. 29 (1981): 4626-4660. EDN: BXJMUK.
• Лаптев А.А. 1984. Численность гнездящихся птиц в дубовых и долинных кедрово-широколиственных лесах Лазовского государственного заповедника // Исследования природного комплекса Лазовского заповедника. М.: 41-43.
• Михайлов К.Е. 2014. Различия в заполнении тайги (сплошных массивов бореальных лесов)
мелкими лесными птицами-мигрантами на примерах нескольких «модельных» для севера Приморского края групп видов Passeriformes. Часть 1 // Рус. орнитол. журн. 23 (978): 773-827. EDN: RXCEAD.
• Михайлов К.Е., Коблик Е.А. 2013. Характер распространения птиц в таёжно-лесной области севера Уссурийского края (бассейны рек Бикин и Хор) на рубеже XX и XXI столетий (1990-2001 годы) // Рус. орнитол. журн. 22 (885): 1477-1487. EDN: QBDPIL.
• Михайлов К.Е., Шибнев Ю.Б., Коблик Е.А. 1998. Гнездящиеся птицы бассейна Бикина (аннотированный список видов) // Рус. орнитол. журн. 7 (46): 3-19. EDN: KTNORV.
Назаренко А.А. (1971а) 2023. Краткий обзор птиц заповедника «Кедровая Падь» // Рус. орнитол. журн. 32 (2333): 3579-3631. EDN: QVHDNF.
• Назаренко А.А. 1971б. Летняя орнитофауна высокогорного пояса южного Сихотэ-Алиня // Экология и фауна птиц юга Дальнего Востока. Владивосток: 99-126.
• Назаренко А.А. 1979. О птицах высокогорий Сихотэ-Алиня // Биология птиц юга Дальнего Востока СССР. Владивосток: 3-15.
• Назаренко А.А. 1984. Птичье население смешанных и темнохвойных лесов Южного Приморья, 1962-1971 гг. // Фаунистика и биология птиц юга Дальнего Востока. Владивосток: 60-70.
• Назаренко А.А. 2014. Новое о гнездящихся птицах юго-западного Приморья: неопубликованные материалы прежних лет об орнитофауне Шуфанского (Борисовского) плато // Рус. орнитол. журн. 23 (1051): 2953-2972. EDN: QWKYLR.
• Назаров Ю.Н. (2001) 2018. Распределение наземных гнездящихся птиц на островах Дальневосточного морского заповедника // Рус. орнитол. журн. 27 (1669): 4561-4569. EDN: UZEPVW.
• Назаров Ю.Н. 2004. Птицы города Владивостока и его окрестностей. Владивосток: 1-276.
• Назаров Ю.Н., Казыханова М.Г., Катрич Л.Н. 1979. Летнее питание овсянок на юге Приморья // Биология птиц юга Дальнего Востока CCCР. Владивосток: 43-55.
• Назаров Ю.Н., Шибаев Ю.В. (1984) 2022. Список птиц Дальневосточного государственного
морского заповедника // Рус. орнитол. журн. 31 (2212): 3329-3349. EDN: NODKXK.
• Начаркин Г.А., Говорова Е.А., Сутырина С.В. 2018. Результаты орнитологических исследований в лесах и лугах Сихотэ-Алинского заповедника в 2017 году // Рус. орнитол. журн. 27 (1565): 613-634. EDN: YLZICW.
• Нечаев В.А., Курдюков А.Б., Харченко В.А. 2003. Птицы // Позвоночные животные Уссурийского государственного заповедника. Аннотированный список видов. Владивосток: 31-71.
• Омелько М.А. 1956. О перелётах птиц на полуострове Де-Фриза // Тр. ДВФ АН СССР 3, 6: 337-357.
• Паевский В.А. 1985. Демография птиц // Тр. Зоол. ин-та АН СССР 125: 1-285.
• Панов Е.Н. 1973. Птицы Южного Приморья (фауна, биология и поведение). Новосибирск: 1-376.
• Пукинский Ю.Б. 1978. О редких и малоизученных птицах бассейна реки Бикин // Природа 1: 56-76.
• Пукинский Ю.Б. 2003. Гнездовая жизнь птиц бассейна реки Бикин // Тр. С.-Петерб. общ-ва
естествоиспыт. Сер. 4. 86: 1-267.
• Романов А.Л., Романова А.И. 1959. Птичье яйцо. М.: 1-620.
• Сотников В.Н. 2023. Каталог коллекций. Птицы – Aves. Оологическая и нидологическая коллекции. Киров, 2: 1-304.
• Спангенберг Е.П. (1965) 2014. Птицы бассейна реки Имана // Рус. орнитол. журн. 23 1065): 3383-3473. EDN: SYCTWJ.
• Тиунов И.М. 2004. Численность и распространение наземных гнездящихся птиц островов
Римского-Корсакова // Дальневосточный морской биосферный заповедник. Биота. Т. 2. Гл. 5. Биота островов: распределение, состав и структура. Птицы островов Римского-Корсакова. Владивосток: 723-758.
• Черский А.И. 1915. Орнитологическая коллекция музея общества изучения Амурского края во Владивостоке // Зап. Общ-ва изучения Амурского края 15: 143-276.
• Шибнев Ю.Б. 1984. Фенологические наблюдения за птицами в заповеднике «Кедровая падь» // Фенологические явления в Приморье. Владивосток: 86-92.
• Шохрин В.П. 2008. Соколообразные (Falconiformes) и совообразные (Strigiformes) Южного
Сихотэ-Алиня. Дис. … канд. биол. наук. Владивосток: 1-205 (рукопись).
• Шохрин В.П. 2017. Птицы Лазовского заповедника и сопредельных территорий. Лазо: 1-648.
• Шульпин Л.М. 1927. Новые данные по распространению птиц в Южно-Уссурийском крае и описание новых форм // Ежегодник Зоол. музея АН СССР 28, 3: 398-406.
• Gamova T.V. 2002. Breeding biology of the Tristram’s bunting Emberiza tristrami in Southern Primorye (Russian Far East) // Avian Ecol. Behav. 9: 23-38.
• Hoyt D.F. 1979. Practical methods of estimating volume and fresh weight of bird eggs // Auk 96: 73-77.
• Nartshuk E.P., Matyukhin A.V., Shokhrin V.P. 2023. Birds as hosts of parasitic louse flies (Diptera) in the south of the Russian Far East // Зоол. журн. 102, 3: 310-316.
Юрий Николаевич Глущенко, Дмитрий Вячеславович Коробов. Тихоокеанский институт географии ДВО РАН, Владивосток, Россия. E-mail: yu.gluschenko@mail.ru; dv.korobov@mail.ru
Татьяна Владимировна Гамова. ФНЦ биоразнообразия наземной биоты Восточной Азии ДВО РАН, Владивосток, Россия. E-mail: birdsdv@mail.ru
Валерий Павлинович Шохрин. Объединённая дирекция Лазовского государственного природного заповедника им. Л.Г.Капланова и национального парка «Зов тигра», с. Лазо, Приморский край, Россия. E-mail: shokhrin@mail.ru
Геннадий Николаевич Бачурин. Научно-практический центр биоразнообразия, Ирбит, Свердловская область, Россия. E-mail: ur.bagenik@mail.ru
Иван Михайлович Тиунов. ФНЦ биоразнообразия наземной биоты Восточной Азии ДВО РАН, Владивосток, Россия. Государственный природный биосферный заповедник «Ханкайский», Спасск-Дальний, Приморский край, Россия. E-mail: ovsianka11@yandex.ru
Николай Николаевич Балацкий. Новосибирск, Россия. E-mail: nnbal54@mail.ru
Дмитрий Анатольевич Беляев. Приморский государственный аграрно-технологический университет, Уссурийск, Приморский край, Россия. Объединённая дирекция государственного природного биосферного заповедника «Кедровая падь» и национального парка «Земля леопарда» им. Н.Н. Воронцова, Владивосток, Россия. E-mail: d_belyaev@mail.ru
Владимир Несторович Сотников. Государственный природный заповедник «Нургуш», Киров, Россия. E-mail: sotnikovkgzm@gmail.com
Источник
Русский орнитологический журнал 2025, экспресс-выпуск 2505. стр. 869-900.