

Статус. Желтогорлая овсянка Cristemberiza elegans (Temminck, 1836) (рис. 1) является обычным, а местами многочисленным пролётным и гнездящимся перелётным видом Приморского края; в небольшом числе зимует в его южной половине, чаще в прибрежной зоне. На территории Приморья обитает подвид C. e. elegans (Temminck, 1836).
Распространение и численность. При наличии подходящих стаций желтогорлые овсянки гнездятся на всей территории Приморского края (Глущенко и др. 2016), включая многие острова, расположенные в заливе Петра Великого (Лабзюк и др. 1971; Назаров, Шибаев 1984; Назаров 2001, 2004; Тиунов 2004; и др.).
По нашим данным, на юге Приморского края, в заповеднике «Кедровая падь», в мае – начале июня наиболее высокую плотность населения отметили в светлых ясенево-ильмовых лесах (достигает 42 пар/км2) и разнотравно-кустарниковых ольшаниках среднего и нижнего уровней реки Кедровая (49-53 пар/км2). В июне-июле численность этих овсянок возрастает на пологих и среднекрутых склонах южных экспозиций с высокотравными и леспедецевыми дубняками (29-32 пар/км2). Гораздо реже (до 6 пар/км2) птицы встречались на сильно осветлённых участках по нижней части пологих склонов, занятых папоротниковыми дубняками.
По материалам А.Б. Курдюкова (2014), в 2008 году в заповеднике и его окрестностях обилие желтогорлых овсянок в разных вариантах широколиственных лесов горных склонов находилось в пределах от 27.1 до 84.0 пар на 1 км2, в хвойно-широколиственных лесах – от 19.9 до 29.6, в долинных лесах – от 23.5 до 69.7, в различных местообитаниях пирогенного древесно-кустарниково-лугового комплекса – от полного отсутствия до 48.0 пар/км2, а в посёлке Барабаш составило 3.4 пар/км2. В национальном парке «Земля леопарда», в долине реки Грязная это многочисенный гнездящийся вид, средняя плотность населения которого в середине апреля и в середине мая варьировала от 30.4 до 20.2 ос./км2 (Беляев и др. 2019).

В 1962-1971 годах в чернопихтово-широколиственных лесах заповедника «Кедровая падь», в хвойно-широколиственных лесах Уссурийского заповедника и в переходных (от смешанных к темнохвойным) лесах истоков реки Уссури в разные годы плотность населения этих овсянок составляла 1.0-2.0; 0.6-5.0 и 0-3.0 пар/км2, соответственно (Назаренко 1984). По другим данным, в Уссурийском заповеднике численность желтогорлых овсянок в разные годы в хвойно-широколиственных лесах варьировала от 0.6 до 10.5, а в широколиственных – от 41.0 до 54.0 пар/км2.
В 2001 году в пойменных кедрово-ильмовых лесах среднем насчитывали 87.1 пар/км2 этих овсянок (Нечаев и др. 2003). В долинных лиственных лесах и дубняках Уссурийского заповедника В.А. Харченко (2015) относила желтогорлую овсянку к числу лидирующих по численности видов птиц, а их обилие в этих биотопах достигало 66 и 10 пар/км2, соответственно. В 2013-2016 годах в девственных неморальных хвойно-широколиственных лесах этого заповедника насчитывали от 1.35 до 7.46 пар/км2 (Курдюков 2017).
По нашим данным, в Уссурийском городском округе в смешанных лесах долин рек Барсуковка и Комаровка плотность гнездования была 1-3 пар/га. В 2002-2005 годах в сопочных дубняках окрестностей Уссурийска относительная численность этих овсянок в летнее время достигала 50 ос./км2 (Глущенко и др. 2006а). На территории лесного участка Приморского ГАТУ за период наблюдений с 2019 по 2022 год желтогорлая овсянка входила в число доминирующих видов птиц (Беляев 2022).
Весной 2019 года плотность её населения в кедрово-широколиственных лесах составляла 27.6 ос./км2 (Беляев 2019); весной 2020 года она колебалась от 23.5 ос./км2 в кедровошироколиственных лесах до 74.8 ос./км2 в долинных широколиственных лесах. Весной 2021 года этот показатель варьировал от 6.0 в кедрово-широколиственных лесах до 66.7 ос./км2 в долинных лесах. В 2023 году в широколиственных лесах долины Комаровки относительная численность желтогорлой овсянки составляла 24.4, а в дубняках в окрестностях села Раковка – 30.0 ос./км2 (наши данные).
В конце 1970-х годов в западных отрогах Сихотэ-Алиня (хребет Синий) в елово-кедрово-широколиственных, в кедрово-широколиственных и в широколиственных лесах относительная численность желтогорлых овсянок составляла 5.1-30.8; 9.4-28.7 и 25.7-53.0 ос./км2, соответственно (Кушнарёв 1984). В 2002-2003 годах на Приханкайской низменности этих овсянок обнаружили локально гнездящимися в дубняках на сопках Гайворонская и Лузанова, а также в пойменных лесах по реке Спасовка, где плотность их населения варьировала от 1.4 до 14.0 пар/км2 (Глущенко и др. 2006б).
В Лазовском заповеднике желтогорлые овсянки относятся к многочисленным птицам, где в 1974-1975 годах в долинных кедрово-широколиственных лесах их обилие составляло 12.5 пар/км2 и 5 % от общей численности птиц, а в дубняках – 8.8 пар/км2 и 8.8 %, соответственно (Лаптев 1984). В дубовых лесах в 1978 году плотность населения этих овсянок не превышала 1.8 ос./км2, в 1988 году достигала 21 пар/км2; в 1994 году – 9.5 пар/км2. В долинных кедрово-широколиственных лесах в 1988 году их относительная численность составляла 32.3 пар/км2, в 1992 – 11.9 пар/км2, а в долинных многопородных лесах в 1993 году – 21.7; в 1994 – 16.7 пар/км2. В 2001 году в долине реки Перекатная плотность населения желтогорлых овсянок была около 80 ос./км2 (Шохрин 2017).
В дубовых лесах на склонах и гребнях гор Среднего Сихотэ-Алиня плотность населения гнездящихся птиц достигала 11 пар/км2; в осиново-берёзовых лесах – 44.4 пар/км2; в кедрово-широколиственных лесах – не более 3 пар/км2 (Кулешова 1969).
В национальном парке «Удэгейская легенда» в среднем течении реки Большая Уссурка желтогорлые овсянки доминировали в долинных кедровниках, где в начале июня 2021 года плотность их населения достигала 52.5 ос./км2, а также в дубняках – 47.1 ос./км2 (Беляев, Коваленко 2023). Во второй половине июля 2020 года эти овсянки встречались здесь с частотой 0.49 ос./км, при этом в основном отмечали нераспавшиеся выводки (Беляев 2022).
На севере Приморья, в долине реки Бикин, желтогорлые овсянки распространены по всему бассейну, при этом в её среднем течении они многочисленны (Пукинский 2003). По другим данным, эти овсянки обычны вверх до села Охотничий, а выше они распространены локально, достигая устья реки Зева (Михайлов и др. 1998). Плотность их населения на Бикине в оптимальных местообитаниях достигает 40-45 пар/км2; реже они встречаются в хвойных и белоберёзовых лесах (не более 12-16 пар на 1 км2), а в целом в этом регионе птицы тяготеют к осветлённым древостоям (Ильинский 1980). На северо-востоке края в различных типах дубняков в 1970, 1974 и 1986 годах плотность гнездящихся желтогорлых овсянок менялась незначительно: 2.0, 4.0 и 4.3 пар/км2, соответственно (Елсуков 1990).
Местообитания. Желтогорлая овсянка является типичным лесным видом, наиболее обычным в долинных ясенево-ильмовых широколиственных лесах, где выбирает для гнездования участки с редким подлеском из кустарников (особенно рябинника рябинолистного Sorbaria sorbifolia) и травянистым покровом из хвоща зимующего Equisеtum hyemаle и страусника обыкновенного Matteuccia struthiopteris. Эти овсянки также селятся в дубняках по южным склонам сопок, достигая высоты 700 м над уровнем моря.
В Южном Приморье типичными гнездовыми биотопами служат дубняки паркового типа, почти лишённые кустарников, а также светлые одноярусные леса из ясеня, бархата амурского и маакии, но отдельные пары гнездятся и в многоярусных лесах с сильным затемнением, но не проникают в горные чернопихтово- и кедрово-широколиственные леса и на участки сильно изменённой урёмы по нижнему течению крупных рек (Панов 1973).
По нашим данным, на юге края, в долине реки Кедровая, желтогорлая овсянка – обычный и местами многочисленный вид, занимающий практически все типы леса, за исключением тёмных пихтово- и кедровошироколиственных лесов верхнего течения. В Уссурийском городском округе, в долинах рек Барсуковка и Комаровка (окрестности села Каймановка), птицы предпочитают разреженные смешанные леса из берёзы, ольхи и ясеня со среднегустым подлеском из рябинника рябинолистного и спиреи иволистной Spiraеa salicifolia и травянистым покровом. На Борисовском плато типичными гнездовыми стациями являются дубовые леса, в том числе светлые дубняки с лиственницей, при этом верхний уровень обитания составляет 550-600 м н.у.м. (Назаренко 2014).
По данным Е.П. Спангенберга (1940), в низовье реки Большая Уссурка (Иман) в 1938 году желтогорлые овсянки заселяли только разреженные леса сопок. На севере края, в таёжной области средне-верхнего Бикина, они сильно привязаны к галерее пойменного леса, который птицы заполняют неровно, с перерывами, образуя, как и в сопочном ландшафте, рыхлые гнездовые парцеллы из 3-5 пар в солнечных парковых микровыделах (Михайлов 2014). По другим данным, в мае желтогорлые овсянки в основном гнездятся в пойменных лесах, а в июнеиюле, когда травянистый покров этих лесов становится слишком высоким, большинство размножающихся особей перемещаются в светлые дубравы ближайших склонов сопок (Пукинский 1974). В верхнем течении Бикина этот вид – немногочисленный, спорадически распространённый обитатель вторичных смешанных лесов с преобладанием лиственных пород (Волков 1997).
По мнению Л.О. Белопольского (1950), в Лазовском заповеднике в выборе мест обитания в гнездовой период желтогорлые овсянки отдают предпочтение пойменным зарослям с большой примесью ольхи и лугам с богатым травостоем. По сведениям, полученным Н.М. Литвиненко и Ю.В. Шибаевым (1971), излюбленными местами гнездования этих овсянок здесь являются вторичные леса, особенно дубняки, растущие на сопках, а также небольшие полянки и вырубки в широколиственных лесах среднего течения реки Киевка. По данным С.В. Винтера и А.И. Мысленкова (2011), эти птицы на заповедной территории обычны и гнездятся на разреженных участках или опушках широколиственных лесов. В северо-восточном секторе Приморья желтогорлая овсянка является обычным видом приморских дубняков, а также долинных широколиственных и смешанных лесов (Елсуков 1999).
Некоторые варианты типичных гнездовых биотопов желтогорлых овсянок иллюстрирует рисунок 2.
Весенний пролёт. В южной половине Приморского края первых прилетевших особей желтогорлой овсянки обычно наблюдали с середины марта, однако в прибрежных районах их нередко регистрировали и раньше, хотя отличить мигрантов от зимовавших в регионе особей не всегда возможно.


В Южном Приморье весенний пролёт наблюдали с первой декады марта по третью декаду апреля (Панов 1973). В окрестностях Уссурийска первое появление явно мигрирующих групп мы отмечали 15 марта 2007, 19 марта 2023, 21 марта 2003, 23 марта 2002, 26 марта 2004, 30 марта 2024 и 31 марта 2006. Пролёт здесь протекает весь апрель и обычно заканчивается в первой декаде мая, но наиболее интенсивная миграция идёт в первой половине апреля, когда нередко отмечали группы, содержащие до 70 особей, а за день наблюдений учитывали более 300 птиц (Глущенко и др. 2006а).
На Приханкайской низменности весной первых желтогорлых овсянок мы регистрировали 15 марта 2002, 23 марта 1973, 28 марта 1972 и 1993, 2 апреля 2003 и 4 апреля 1978. Массовый пролёт здесь проходит во второй и третьей декадах апреля, однако ещё и в первой декаде мая в несвойственной для размножения обстановке эти овсянки ещё вполне обычны. Последние весенние встречи явно пролётных птиц здесь отметили 21 мая 1972 и 23 мая 1980 (Глущенко и др. 2006б).
В Лазовском заповеднике и его окрестностях в середине ХХ столетия первое весеннее появление зарегистрировали 4 апреля 1944 и 8 апреля 1945, а к концу этого месяца массовый пролёт заканчивался (Белопольский 1950). В 1982 году первые самцы появились здесь 25 марта (Шохрин 2017). По нашим данным, в XXI столетии в окрестностях заповедника самые ранние даты прилёта желтогорлых овсянок приходились на 15 марта 2009, 16 марта 2011, 17 марта 2001 и 2004; средняя многолетняя дата появления – 25 марта, а наиболее поздние из первых весенних регистраций датированы 1 апреля 1985 и 1996.
В районе залива Восток весенняя миграция проходит в третьей декаде марта и в апреле (Нечаев 2014). На островах залива Петра Великого мигрирующие желтогорлые овсянки многочисленны в апреле, а последних пролётных птиц здесь отмечали в первой декаде мая (Лабзюк и др. 1971). В целом в настоящее время в Южном Приморье транзитные особи и группы желтогорлых овсянок чаще всего встречаются во второй половине марта и в первой половине апреля (рис. 3).
Гнездование. Местные птицы занимают гнездовые участки и разбиваются на пары ещё до того, как заканчивается миграция. Одиночных самцов, занявших территорию, на юге Приморья наблюдали 16 апреля 1962, а сформировавшиеся пары неоднократно попадались 20 апреля 1960 и 21 апреля 1962 (Панов 1973). В бассейне реки Бикин поющих птиц отмечали со второй декады апреля; с середины мая регулярное пение прекращалось, но во второй декаде июня оно возобновлялось и к концу этого месяца вновь становилось обычным (Пукинский 2003).
По нашим данным, с момента прилёта до начала насиживания самцы активно поют регулярно, особенно до 13 ч и в 15-19 ч. Можно слышать два типа песен: громкие полные с большими межпесенными интервалами (повседневные), исполняемые на деревьях на высоте до 3 м, и тихие неполные с короткими межпесенными интервалами (территориальные) – на дереве ниже или на земле. Частота пения варьирует от 4 до 20 песен в 1 мин, а межпесенные интервалы длятся 2-22 с. Дневные пики активности пения отмечены в 12, 17 и 19 ч. Пение обычно стихает с середины третьей декады июня.
Границы индивидуальных участков нечёткие и перекрываются у разных пар. Площадь охраняемой территории варьирует от 1600 до 5500 м2. Разные пары могут кормиться всего в нескольких сантиметрах друг от друга, без проявления агрессии. Копуляцию у овсянок отмечали за 2-3 дня до строительства гнезда, и этот процесс периодически повторяется до четвёртого дня насиживания кладки. Самка свистом даёт понять самцу, находящемуся в 3-5 м от неё, что готова к копуляции. Кормящийся самец, издавая тихую непрерывную песню, начинает гоняться за самкой. Догоняет её на земле или невысоком кустике и совершает садку в течение 5-10 с. Самка издаёт двухсложное «чру-чру». После этого самец отлетает, а самка остаётся на том же месте и продолжает кормиться. Затем она садится на дерево или другое растение и совершает смещённые чистки оперения груди и хвоста.
В течение всего гнездового сезона партнёры кормятся вместе, в 0.3-5.0 м друг от друга. Во время кормёжки самец поддерживает связь с самкой, изредка подавая тихую позывку «тик-тик». Желтогорлые овсянки активно поют в периоды насиживания и выкармливания птенцов. Песенные посты находятся вблизи гнезда, в 30-60 м от него, в центре гнездового участка. Покормившись, самец поёт недалеко от места кормёжки 4-5 тихих песен с короткими межпесенными интервалами, а через несколько минут на своём постоянном песенном посту выдаёт более громкие обычные песни. Во время насиживания самцы становятся агрессивными и не терпят близкого соседства других самцов. Услышав пение соседнего самца рядом со своим песенным постом, хозяин участка начинает гоняться за ним по кругу или зигзагообразной траектории. В ответ на пение конкурента он поёт сначала тихо, а потом громче. Соседний самец при этом меняет обычный вокальный стиль на серию непрерывных территориальных песен, а после окончания пения он часто остаётся покормиться на чужом участке. Самец-хозяин оставляет его в покое и поёт в обычном песенном стиле, но гораздо громче.
Обычно птицы исполняют песни, сидя сравнительно открыто на возвышенном месте (рис. 4).

Гнездовой период длится около трёх месяцев: с третьей декады апреля по середину июля (табл. 1), за который многие пары успевают вырастить два выводка.
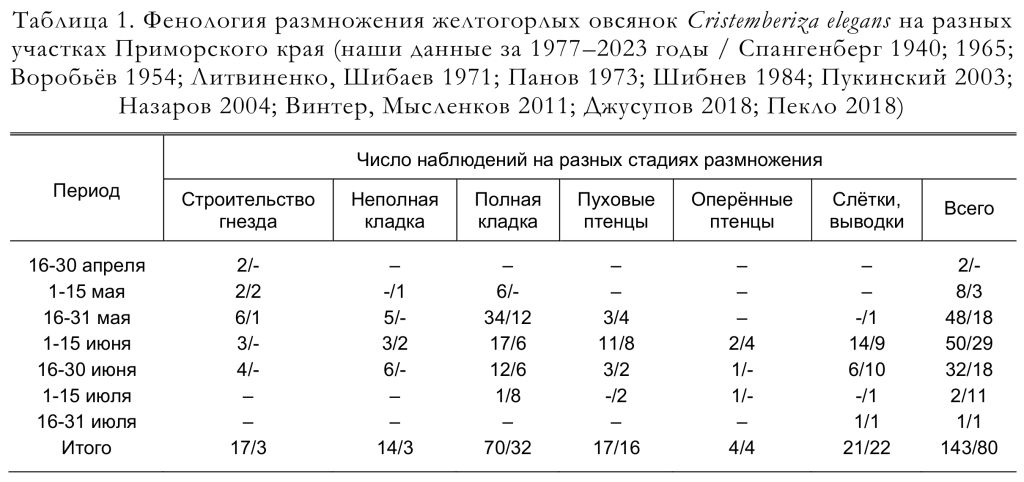
В 1972-1975 годах в Южном Приморье массовое строительство гнёзд наблюдали с 5 по 9 июня (данные Ю.Н. Назарова). Здесь же в 1997 году этот процесс проходил 21-24 мая, 11-16 июня, а у некоторых пар он затянулся до 27 июня; в 1998 – 25-29 апреля, 3-22 мая и 12 июня; в 1999 – 20-23 мая (наши данные). В бассейне реки Бикин строительство гнёзд отмечали в мае и июне (Пукинский 2003).
Строительство первых гнёзд занимает 3-4 дня, а последующих может сокращаться на один день. Работа начинается в 6 ч 40 мин, заканчивается после 18 ч, а наиболее интенсивно она протекает с 7 ч 00 мин до 11 ч 30 мин и с 14 ч 20 мин до 18 ч 00 мин. Самка прилетает к гнезду со строительным материалом через 2-5 мин, что за 1 ч составляет 7-13 прилётов. Укладка одной порции принесённого материала занимает 40-50 с. Во время строительства партнёры держатся рядом: в 74 % случаев они улетают вместе покормиться, а возвращаются вдвоём только в 32 % наблюдаемых ситуаций. Птицы добывают пищу в 100-120 м от гнезда.
Гнёзда, найденные К.А. Воробьёвым (1954) в Южном Приморье 22 мая 1948 и 5 июня 1949, размещались на земле. Гнёзда, осмотренные Е.Н. Пановым (1973) 21 мая 1962, располагались в основании дерновин злаков и одно из них было хорошо закрыто сверху. Гнездо, обнаруженное 23 мая, было свито под прикрытием сухой палки и куртины травы (Панов 1973). По данным С.В. Винтера и А.И. Мысленкова (2011), в районе Лазовского заповедника желтогорлые овсянки гнездились на разреженных участках или опушках широколиственных лесов. Постройки (n = 6) были расположены на обочинах заброшенных лесных дорог (4 случая), у тропы (1) и на поляне среди пойменного редколесья (1); две из них были свиты на земле, одна – на стеблях осоки в 12 см от земли, а ещё три – на кустах на высоте 0.5 м.
В бассейне реки Бикин в ранние сроки птицы гнездятся преимущественно в углублениях почвы, под прикрытием прошлогодней растительности; июньские постройки обычно помещаются в зелёных розетках листьев папоротника, на кустах элеутерококка, бузины или в сплетениях лимонника на высоте 0.2-1.3 м от земли (Пукинский 2003).

По нашим данным, первые гнёзда желтогорлые овсянки строят на земле (среди травы, в розетках папоротника или между побегами кустарников). Более поздние гнёзда птицы нередко приподнимают над поверхностью земли (в среднем на 12 см), укрепляя их в перекрёстах 2-3 веточек растения, которые крепко сжимают или пронизывают стенки. Из 59 описанных нами гнёзд 45 располагались на земле, а остальные были построены на кустарниках на высоте от 7 до 123 см.
Некоторые типичные варианты расположения гнёзд на земле и на древесных растениях иллюстрируют рисунки 5 и 6.

Гнездо, осмотренное К.А. Воробьёвым (1954, с. 187) 22 мая 1948, было сделано «из широкой сухой травы; довольно глубокий лоточек был выложен шерстью косули и конским волосом. Сверху гнездо было хорошо закрыто травой и крапивой». Гнёзда, найденные Е.Н. Пановым (1973), состояли из сухой травы, а их лотки были выложены серой шерстью.
Материалом для гнезда, обнаруженного Ю.Н. Назаровым (2004) 24 июня 1992 в окрестностях Владивостока, служили листья осок, луб, стебли и листья злаков, лист дуба и вайя папоротника, а лоток был выстлан корешками папоротника. Постройка, найденная в дельте реки Раздольная 5 июля 1975, была свита из стеблей подмаренника и ломоноса, а в лотке были корешки папоротника. Материалом для гнезда, осмотренного в долине реки Суходол 23 июня 1972, были крупные листья и куски стеблей злаков, а в лотке находилась шерсть. Постройка, найденная там же 26 июня 1973, состояла из стеблей и листьев злаков и кусков коры, а лоток выстилали тонкие стебельки злаков и небольшое количество шерсти. Гнездо, осмотренное там же 30 июня 1975, было построено из стеблей злаков, кусков коры тополя, а в лотке были тонкие стебельки и немного конских волос (Назаров 2004).
По данным С.В. Винтера и А.И. Мысленкова (2011), в окрестностях Лазовского заповедника материалом для наружного и среднего слоёв гнёзд служили листья и стебли злаков, лыко и кора лещины, а их лотки были выстланы конскими волосами, лишайником рода Usnea, тонкими стеблями злаков, остевыми волосами кабана и енотовидной собаки.
По описанию Ю.Б. Пукинского (2003), размеры и конструкции разных гнёзд довольно изменчивы: наиболее массивными и разнообразными по строительному материалу являются постройки, расположенные на кустах и папоротнике. Они имеют рыхлое основание, состоящее из прошлогодней травы, массивный внешний слой, выстроенный из грубых стеблей и листьев злаков, крапивы и других растений, а также средний слой, сплетённый из полосок луба с добавлением стеблей злаков. Выстилка лотка бывает разной по толщине; она плотно соединяется со средним слоем и состоит из тонких корешков, травинок, конских волос и ости копытных. Гнёзда, построенные на земле, лишены основания и массивного внешнего слоя, представляя собой плотные чашечки, свитые главным образом из луба и выстланные грубой шерстью (Пукинский 2003).
По нашим данным, в весовом отношении в постройках желтогорлых овсянок преобладают древесные компоненты (кора и лубяные волокна деревьев), составляющие до 61-90 % массы гнезда (табл. 2).

Из всех видов овсянок кору деревьев в гнёздах мы находили только у желтогорлой, и этот компонент встречен в 43-64 % построек. Шерсть лошадей и собак присутствовала в 64 % гнёзд, а её весовой вклад в лотке составлял около 11 %. В отличие от других видов овсянок, во внутренней части гнёзд желтогорлой овсянки отсутствуют листья трав, кустарников и деревьев. Их заменяют корни трав и древесные компоненты. Длина травинок и утепляющего материала варьировала от 64 до 522, в среднем составляя 271.7±144.1 мм, а их ширина находилась в пределах от 2 до 24, в среднем 9.7±7.2 мм. Во внешней части гнезда, кроме стеблей и листьев злаков – главных составляющих построек овсянок, в большом количестве встречались древесные компоненты, стебли и листья других трав, а также листья деревьев и кустарников (до 23 %). Длина материала во внешней части гнезда 91-298, в среднем 176.8±56 мм, а ширина 0.5-16, в среднем 6.2±5.2 мм.
В разных случаях лоток гнезда бывает неглубоким, рыхлым и неаккуратным, либо глубоким, рыхлым и аккуратным. Масса лотка варьирует от 1.9 до 9.6, в среднем 6.21 г. Общая средняя масса гнезда составляет 15.26 г. Форма постройки чашевидная, сужающаяся книзу, или асимметричная. Большинство гнёзд асимметричные, при этом их стенки неплотные, а их общий вид выглядит неряшливо. Размеры построек представлены в таблице 3.
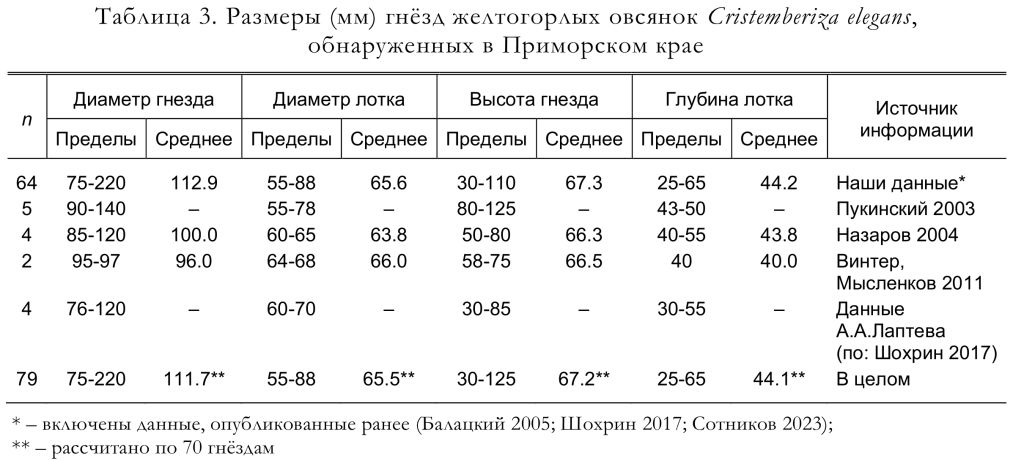
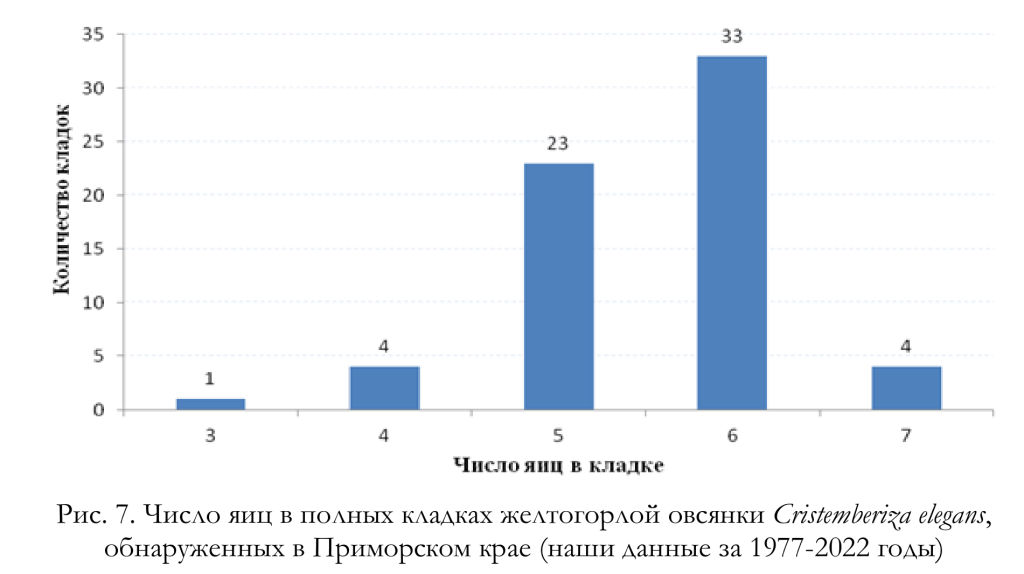
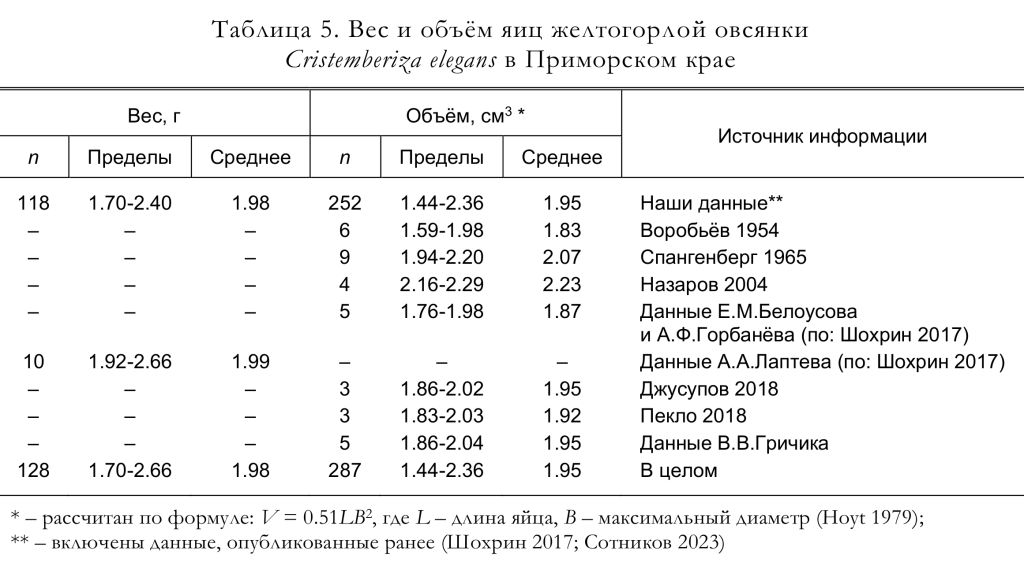
По нашим данным, к откладке яиц самки приступают на следующий день либо через 1-3 дня после окончания строительства гнезда. Полные кладки, осмотренные Ю.Б.Пукинским (2003) в бассейне реки Бикин, состояли из 4 (1 случай), 5 (9) и 6 (3) яиц. По данным Ю.Н. Назарова (2003), найденные гнёзда содержали 4 (1 случай) и 5 (3) яиц. Согласно нашим материалам, число яиц в полных кладках варьировало от 3 до 7, чаще всего их было 6 (рис. 7), а средняя величина кладки составила 5.54 яйца (n = 65). Линейные размеры, индекс удлинённости, вес и объём яиц желтогорлых овсянок приведены в таблицах 4 и 5.
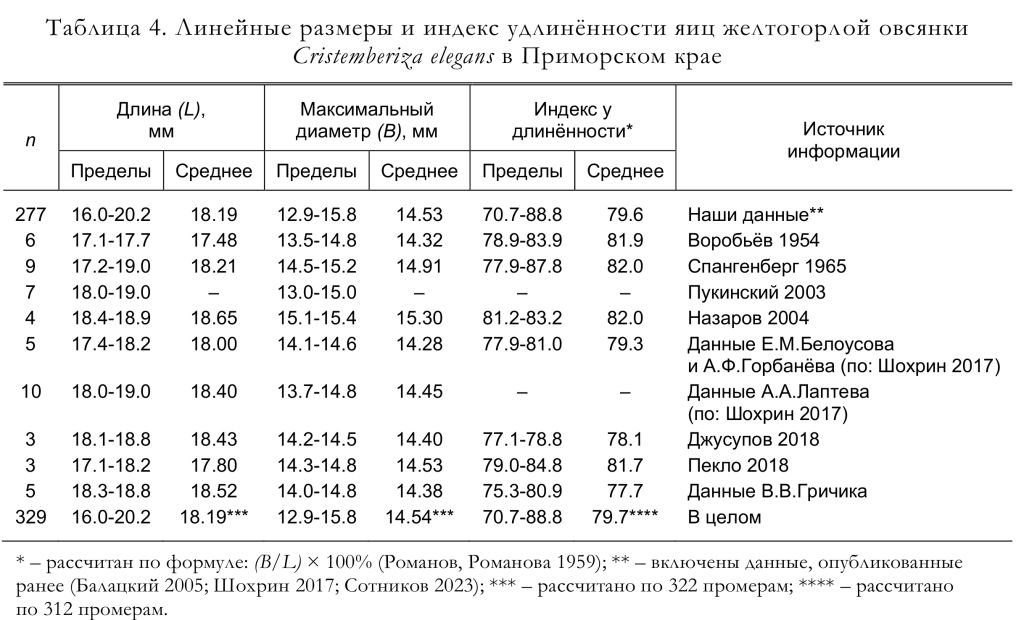
По данным К.А. Воробьёва (1954), яйца имеют белый фон, по которому разбросаны бурые точки и крапинки, гуще сконцентрированные на тупом конце. Согласно описанию Е.П. Спангенберга (1965), скорлупа


яиц блестящая, её основной фон голубовато- или розовато-белый; мелкие точки и пятна разбросаны редко и равномерно, либо собраны в виде венчика на тупом полюсе яйца; внутренние точки светло-серые, а поверхностные – бурые. Ю.Б. Пукинский (2003, с. 229) пишет, что «скорлупа беловатая с лиловым оттенком. Редкий рисунок из небольших пятен (разной величины и неправильной формы) интенсивного лиловато-чёрного цвета; некоторые пятна соединяют “жилки”; у половины яиц рисунок сгущается на тупом полюсе в виде венчика».
По нашим данным, окраска яиц слабо изменчивая. Общий фон яйца от светло-кремового до светло-бурого. Рисунок пятнисто-линейный. Пятна от светло-серых или светло-коричневых до тёмно-бурых и чёрных. Они покрывают яйцо либо равномерно, либо группируются на тупом конце, где часто образуют венчик. У яиц одной или разных кладок плотность пятнистого рисунка может варьировать от редкой до сплошной (иногда занимает до 80 % тупого конца). Пятна поверхностные или глубинные, диаметром 1-3 мм. Линейный рисунок, как правило, редкий, поверхностный и более светлый (оттенки коричневого). Линии простые или разветвлённые, длиной 1-4 мм, диаметром 0.1-1.0 мм. Поверхность яйца со слабым блеском. Для наглядности приводим коллажи, составленные из фотографий различных кладок желтогорлых овсянок из Приморского края, выполненных в природных условиях (рис. 8) и в студии (рис. 9).
По данным Ю.Б. Пукинского (2003), насиживание, в котором принимают участие оба партнёра, продолжается 12 сут (1 наблюдение), при этом в мае самец проводит в гнезде половину дневного времени. Согласно нашим наблюдениям, птицы начинают насиживать после откладки предпоследнего яйца и продолжают в течение 10-12 сут. Степень участия партнёров в насиживании неодинакова: либо в первые дни инкубации самец сменяет самку только на время её кормёжки, а в последующие дни делит эту обязанность с ней наравне, либо их участие паритетное на протяжении всего периода насиживания кладки, при этом партнёры поочерёдно сменяют друг друга. В первые два дня самка сидит на яйцах 10-36 мин и отлучается на 18-23 мин; в последующие дни продолжительность сеанса насиживания возрастает до 70-103 мин, а время отсутствия сокращается до 5-12 мин. Самец однократно насиживает по 43-86 мин и отлучается от гнезда на более длительное, чем самка, время (до 25 мин). Если самец долго не сменяет самку, она садится на край постройки или бегает возле неё, склёвывая насекомых.
С гнезда насиживающие птицы слетают на 2-4 м, сначала в траву, а затем садятся на ближайшее дерево на высоте до 2 м и спустя 10-15 с улетают. В период насиживания партнёры кормятся вместе, в 0.2-5.0 м друг от друга. Самец в это время поддерживает контакт с самкой тонкими отрывистыми позывками. Пищу для птенцов (личинки, имаго пядениц, личинки жуков, перепончатокрылые) овсянки собирают главным образом на земле среди коряг и опавшей листвы. По нашим данным, успешность насиживания варьирует от 25 до 87 %, яйца-болтуны составляют 2.2 % от погибших, а в среднем отход равен 0.6-2.0 яйца на гнездо (наши данные).
По описанию И.А. Нейфельдт (1970), однодневный птенец желтогорлой овсянки имеет пух на 8 птерилиях. По нашим данным, пух присутствует на 10 птерилиях: надглазничных, затылочной, плечевых, локтевых, кистевых, спинной, брюшных, бедренных, голенных и копчиковой.
На копчике пух рудиментарный, из 8 остинок. Цвет пуха варьирует от белого на кистевых, брюшных, голенных и копчиковой птерилиях до светло-серого – тёмно-серого на остальных. Длина пуха максимальна над глазами (6-10, в среднем 8.0 мм), на затылке (8-10, 9.0 мм) и спине (9-10, 9.5 мм), а минимальна – на локтях (0-0.5, 0.3 мм), голенях (0-1.5, 0.8 мм) и копчике (0-1.5, 0.8 мм). На плечах длина пуха 5-7, в среднем 6 мм, на кисти 0.2-4.0, в среднем 2.1 мм, на брюхе 2-5, в среднем 3.5 мм, а на бёдрах – 3-5, в среднем 4 мм. Клюв телесно-серый или буроватый, надклювье буроватое, конёк жёлтый, подклювье жёлтое. Окраска яйцевого зуба варьирует от белого до телесно-серого; клювные валики белые или с жёлтым оттенком. Ротовая полость в разных случаях от беловатой до ярко-розовой. Нёбо ярко-жёлтое или розовато-красное; язык розовый или красный; окраска кожи варьирует от светло-телесной до оранжевой; веки серые; когти белые или телесно-розовые; лапы розовато-оранжевые или светло-жёлтые; ноздри овальные, под углом 45є к коньку клюва.
В возрасте 1 сут птенцы желтогорлых овсянок уже способны издавать тихий писк в ответ на позывки родителей, находящихся в 1.0-1.5 м от гнезда. Этот писк становится громким на 5-е сутки их жизни. Реакция выпрашивания проявляется даже при позывках других видов птиц в 3-4 м от гнезда. На 8-е сутки птенцы при малейшем движении взрослой птицы в гнезде открывают клювы и держат их в таком положении 1-2 мин, высоко приподнимаясь на лапках.
Вес однодневного птенца варьирует от 2.33 до 2.57 г, в среднем составляя 2.45 г (n = 14). Наибольший суточный прирост массы тела происходит на 2-4-е сут (47.5-73.0 %), а минимальный – на 5-6-е сут, когда начинают распускаться кисточки перьев. Оперение (грудные, бедренные, голенные, брюшные, спинные, горловые, плечевые и другие птерилии) интенсивно растёт на 5-е сутки (более 200%). Части тела имеют максимальный прирост на 2-6-е сут (24.0-58.6%). На 2-е сутки глаза приоткрываются на 10%, на 4-е они наполовину открыты, а на 6-е открыты полностью. Слуховые отверстия открываются на 3-е сут. В это же время первыми над кожей появляются пеньки первостепенных и второстепенных маховых перьев. На 5-е сут отрастают пеньки остальных перьев. На 6-е сут распускаются кисточки плечевых, спинных и рулевых перьев. На 9-е сут основные части тела прекращают расти, зато активно распускаются кисточки остальных перьев. Наиболее плавно растут клюв, предплечье, голень, а из перьев – кроющие бедра и первостепенные маховые. На 10-е сут клюв в среднем равен 10.5 мм; кисть 22.0; цевка 23.2; первостепенные маховые 37.0; второстепенные маховые 21.0; рулевые 20.0; теменные 10.0 мм (табл. 6, 7).
У 9-10-дневного слётка клюв становится серовато-коричневым, с более тёмным подклювьем; клювные валики насыщенно-жёлтые; конёк клюва светло-коричневый; цевка розовато-жёлтая; когти светло-серые.
Первостепенные маховые на 7-е сут бурые, на 8-е – тёмно-бурые со светло-жёлтыми каёмками по внешней части опахала, на 9-е сут каёмки становятся светло-рыжими, хорошо видны тёмно-серые пеньки, на 10-11-е сут рыжие каёмки у 1-4 первостепенных маховых остаются только на внутренней части пера. На 10-11 сут у кроющих первостепенных маховых и второстепенных маховых на 1-4 перьях светло-рыжие каёмки на внешней части опахала становятся буровато-серыми, на остальных перьях рыжеватая каёмка имеется на внутренней части опахала. Кроющие второстепенных маховых с 7 по 11 сут остаются тёмно-бурыми с ярко-рыжими каёмками. Верхние кроющие крыла коричневые с жёлтыми пестринами. Теменные из коричневых (на 7-е сутки) становятся серовато-


бурыми с жёлтыми, а затем со светло-коричневыми пестринами на 10-11-е сут. Горловые из кремово-коричневых (на 7-е сут) становятся серовато-коричневыми с тёмно-бурыми, и позже с тёмно-жёлтыми пестринами. На спине коричневый цвет перьев постепенно рыжеет, а пестрины темнеют от белого до тёмно-жёлтого цвета. Бедренные (кремово-серые) перья не претерпевают изменений в цвете, а бледно-кремовые голенные приобретают на 9-е сут светло-жёлтые пестрины. На грязно-белых брюшных перьях к 11 сут появляются светло-бурые пестрины. Копчиковые перья постепенно темнеют от белого до светло-жёлтого. Рулевые темнеют от коричневого до тёмно-бурого с более светлыми краями (наши данные).
Вылупление птенцов мы наблюдали 19 мая 2004 (рис. 10), 23 мая 2002, 27 мая 1995, 1 июня 1989, 11 июня 2015, 13 июня 1999, 14 июня 1998 и 29 июня 1981. Гнездовых птенцов (рис. 11) регистрировали со второй половины мая до середины июля (табл. 1).

Самец и самка участвуют в обогревании птенцов примерно в равной степени (самка до 48-55 мин/ч, самец до 40-50 мин/ч). Наиболее осторожно и скрытно птицы держатся у гнезда с 6-10-дневными птенцами. Перед тем как залететь в него, они выжидают в течение 1 мин в 3 м от него. Незаметно проникают в гнездо и так же незаметно покидают его.
Самец сидит на птенцах по 5.0-6.3 мин, а в случае тревоги становится в полный рост. Покормив птенцов, он дожидается появления самки. Вылетает в ответ на шум её крыльев и тихую тонкую отрывистую 2-4-слоговую позывку, подаваемую в 10-12 м от гнезда. Самка, покормив птенцов, прежде чем начать их обогревать, может сидеть на краю постройки до 10 мин (на 6-е сутки она может находиться в гнезде до 4.5 мин). Через каждые 4 мин она приподнимается на лапках и сотрясающими движениями раздвигает стенки гнезда, освобождая его от отшелушившихся частей трубочек перьев.

При одновременном появлении партнёров у гнезда тот, кто приносит небольшую порцию пищи, отбирает часть корма у другого и кормит ею птенцов. Птица, не покормившая птенцов, съедает оставшихся насекомых и улетает за новой порцией. Самка, проводящая больше времени в гнезде, чаще съедает принесённый корм или вовсе прилетает к птенцам без него. Вылетает из гнезда в ответ на свист самца в 6 м от гнезда. При опасности у гнезда самец адресует самке протяжную позывку, и она продолжает оставаться в нём.
Восьмидневных птенцов родители обогревают в течение 0.7-2.0 мин. 9-дневные птенцы издают позывки, напоминающие таковые взрослых птиц, на которые прилетают родители с кормом, находясь в 30-40 м от гнезда. Корм родители добывают в 6-8 м от гнезда. Интенсивность выкармливания зависит от времени суток и возраста птенцов. 5-дневные птенцы получают от родителей до 8 порций корма за 1 ч, 6-дневные – до 12, 8-дневные – до 13 порций. В течение суток активность выкармливания максимальна в 9-11 и 17-18 ч (в среднем 4.5 посещений), а минимальна в 14-15, 19-20 ч (2.7 посещений). Степень участия родителей в выкармливании неодинакова. До 4-дневного возраста птенцов самка прилетает с кормом до 2.3 раз/ч, а самец 1.2 раз/ч. На 4-8-е сутки самка приносит пищу 3.3 раз/ч, самец 2.4 раз/ч.
Птенцы покидают гнездо на 7-10-е сут. Молодых птиц можно встретить со второй декады июня (рис. 12) до середины августа, когда происходит осенняя линька (со сменой перьев спины, поясницы, нижних кроющих крыла). Первых слётков в 2021 году в долине Большой Уссурки мы встречали 6 июня. В Южном Приморье массовый вылет птенцов из гнёзд в 1972-1998 годы происходил с 10 по 21 июня, и со 2 по 17 июля. В первые три дня после вылета молодые овсянки перемещаются на 6-35 м от гнезда, а ещё через двое суток – на 100-120 м (наши данные).


В Приморье общие потери среди птенцов в 1972-1998 годах составили 10 %, или 0.8 птенца на одно гнездо (данные Ю.Н. Назарова и наши). Слётки одного выводка вследствие неодинаковых темпов роста ведут себя по-разному. Более развитые особи проводят время на нижних ветвях деревьев. При опасности они перелетают на небольшое расстояние, а затем затаиваются, тесно прижав горизонтально расположенное туловище к присаде. Слётки, которые послабее, держатся на низких кустах или на земле, а при тревоге предпочитают затаиваться в траве.
Распад выводков происходит при возрасте молодых в 23-30 дней, то есть спустя 2-3 недели после их вылета из гнёзд, но некоторые выводки продолжают держаться с родителями и позднее. Возле выводка самцы ведут себя более заметно. Часто они располагаются спиной к наблюдателю, поворачивая голову в сторону, при этом их крылья приспущены.
Родители продолжают активно кормить 11-15-дневных слётков. Принося им корм, они подлетают сначала к одному птенцу, а к остальным приближаются незаметно по траве. Самец постоянно следит за окружающей обстановкой, в перерывах между кормлением находится рядом с молодыми, а во время доставки корма совершает короткие высокие разведывательные прыжки по земле. Ещё через неделю взрослые прекращают кормить слётков. Самостоятельных молодых овсянок мы наблюдали со второй половины июня (рис. 13).
Осенние миграции и зимовки. По данным Е.Н. Панова (1973), в Южном Приморье в июле и августе желтогорлые овсянки держатся небольшими одновидовыми стайками, обычно в свойственных им местообитаниях, широкие кочёвки начинаются с начала сентября, а массовый пролёт происходит в октябре (Панов 1973). По нашим данным, в конце июля – августе стайки желтогорлых овсянок многочисленны среди полынных, осоково-разнотравных зарослей, разреженных ясеневников с примесью ильма и черёмухи. В конце июля молодые самцы пробуют голос, сидя высоко на дереве (до 12 м). Их песня звучит тихо, продолжительно, без межпесенных интервалов. В августе стайки, состоящие из молодых из 2-3 выводков и взрослых птиц, держатся высоко на деревьях или среди редких кустарников, а кормятся на земле на открытых участках. В сентябре-октябре мигрирующих желтогорлых овсянок можно наблюдать в несвойственных им стациях, например, в безлесной долине реки Раздольная среди полей с редкими деревцами и кустарниками.
В районе залива Восток миграция желтогорлых овсянок проходит с сентября по начало ноября (Нечаев 2014). На островах залива Петра Великого осенний пролёт начинается в последних числах сентября (Лабзюк и др. 1971). В Лазовском заповеднике в разные годы миграция протекала примерно одинаково: интенсивный пролёт проходил в третьей декаде сентября и в первой декаде октября; во второй декаде октября пролёт ослабевал, а далее сходил на нет. В некоторые годы, например 2015, ещё один пик пролёта бывает во второй декаде сентября (Шохрин 2017). По результатам массового отлова птиц паутинными сетями, проводимого в окрестностях Лазовского заповедника в 2001-2013 годах, начало осенней миграции отметили 2 сентября (2005), а её окончание – 15 ноября (2005) (Шохрин 2014). В некоторые годы в июле, августе и начале сентября желтогорлых овсянок наблюдали в кочующих смешанных стайках вместе с синицами и пеночками.
В Уссурийском заповеднике осенний пролёт желтогорлых овсянок продолжается до середины или до конца ноября, а в отдельные годы эти птицы здесь зимуют (Харченко 2002). Наиболее поздние встречи желтогорлых овсянок в разных районах Приморского края обычно датированы разными числами ноября, реже последней декадой октября либо началом декабря (табл. 8), хотя в последнем случае уже можно говорить о встречах зимующих особей.

В юго-западном Приморье, в заповеднике «Кедровая падь», нерегулярно и в небольшом количестве (1-8 особей) желтогорлые овсянки зимуют в долине реки Кедровая (Назаренко 1963); в другой публикации указано, что зимующие птицы здесь связаны с культурным ландшафтом (Назаренко 1971). О.А. Бурковский (устн. сообщ.) встречал этих овсянок на огородах около посёлка Заводской (микрорайон города Артём) 6 февраля 1996.
В окрестностях Лазовского заповедника желтогорлые овсянки, оставшиеся на зимовку, держались в окрестностях сёл стайками из нескольких птиц (Литвиненко, Шибаев 1971). По данным А.А. Лаптева (письм. сообщ.), одиночных особей здесь наблюдали 2 декабря 1974, 4 января 1974 и 17 февраля 1974. Одиночного самца встретили 10 декабря 1981 на окраине посёлка Преображение, а в окрестностях села Лазо желтогорлых овсянок отмечали 10 января 1999 (2 особи), 18 января 1999 (1), 15 января 2004 (4), 4 декабря 2006 (3), 12-17 января 2007 (3), 9 февраля 2007 (1 особь) (Шохрин 2017), 5 января 2018 (рис. 14.2) и 18 января 2020 (рис. 14.4).

В окрестностях Находки самцов встречали 9 февраля 2019 (рис. 14.9) и 1 января 2023 (рис. 14.1); в Шкотовском районе самца наблюдали 5 февраля 2023 (рис. 14.5); в окрестностях села Корсаковка Уссурийского городского округа самца сфотографировали 14 января 2024 (рис. 14.3).
На Приханкайской низменности одну зимующую особь отметили 24 декабря 1985, а в окрестностях Уссурийска зимовку одиночных особей и группы, насчитывающие до 5 птиц, регистрировали в декабре 2002, а также в январе и феврале 2003 года (Глущенко и др. 2006а).
Питание. Взрослые желтогорлые овсянки собирают корм на земле, часто среди опавших листьев, на ветвях и листьях деревьев (Назаров и др. 1979). Они могут подвешиваться к кончикам ветвей или склёвывать с них добычу на лету (Панов 1973). Животная пища довольно однообразна (табл. 9) и сходна у взрослых и птенцов (Назаров и др. 1979). При поимке крупных гусениц птицы их умерщвляли и выдавливали часть содержимого (рис. 15).


Неблагоприятные факторы, враги, гибель. Основными врагами желтогорлых овсянок являются большеклювые вороны Corvus macrorhynchos. Помимо этого, яйца овсянок съедают змеи (Ю.Н. Назаров, устн. сообщ.; наши данные). На острове Большой Пелис в заливе Петра Великого желтогорлую овсянку (1 экз.) в 1979-1980 годах отметили в питании сапсана Falco peregrinus (Назаров, Трухин 1985). В окрестностях Лазовского заповедника желтогорлых овсянок встречали в добыче перепелятника Accipiter nisus, малого перепелятника A. gularis и ошейниковой совки Otus bakkamoena (Шохрин 2008; 2017; наши данные).
При осмотре более чем 80 гнёзд желтогорлых овсянок, содержащих кладки, нами только однажды, 26 июня 2000 в окрестностях села Красный Яр (Пожарский район), обнаружено яйцо обыкновенной кукушки Cuculus canorus. Оно заметно отличалось от всех известных нам яиц этого гнездового паразита своеобразным фоном и рисунком на скорлупе (рис. 16), больше напоминающим таковые не желтогорлой, а таёжной овсянки Ocyris tristrami.

В 2017-2023 годах при отловах птиц установили, что желтогорлые овсянки являются хозяевами 4 видов мух-кровососок (Hippoboscidae, Diptera): Ornithoica momiyamai (сняли 84 особей), Ornithoica unicolor (18), Ornithomya avicularia (18) и Lipoptena fortisetosa (2) (Nartshuk et al. 2022, 2023; наши данные).
Трёх желтогорлых овсянок, сбитых автомобилями на трассе Лазо – Сергеевка, нашли 15 мая 2018, 2 июля 2020 и 13 сентября 2022.
Литература
• Балацкий Н.Н. 2005. К авифауне верхнего течения Бикина // Рус. орнитол. журн. 14 (278): 98-103. EDN: IJVUSN
• Белопольский Л.О. 1950. Птицы Судзухинского заповедника (воробьиные и ракшеобразные) // Памяти академика П.П. Сушкина. М.; Л.: 360-406.
• Беляев Д.А. 2019. Весенняя орнитофауна южной части лесного участка Приморской ГСХА (Южное Приморье) // Роль аграрной науки в развитии лесного и сельского хозяйства Дальнего Востока. Уссурийск: 161-170.
• Беляев Д.А. 2022а. Биоразнообразие орнитофауны лесного участка Приморской ГСХА (Южное Приморье) // Гуковские чтения. Уссурийск: 21-26.
• Беляев Д.А. 2022б. Предварительные данные о населении птиц бассейна реки Большая Уссурка (Национальный парк «Удэгейская легенда», Приморский край) // Вестн. ИрГСХА 3 (110): 45-63.
• Беляев Д.А., Глущенко Ю.Н., Коробов Д.В., Тиунов И.М. 2019. Птицы бассейна верхнего течения р. Грязная (национальный парк «Земля леопарда») // Биота и среда заповедных территорий 4: 65-85.
• Беляев Д.А., Коваленко Д.Б. 2023. Данные о населении птиц бассейна реки Большая Уссурка (национальный парк «Удэгейская легенда») по результатам учётов 2021 года // Лесное хозяйство: материалы 87-й науч.-техн. конф. Минск: 49-53.
• Винтер С.В., Мысленков А.И. 2011. О птицах Лазовского заповедника // Сомовская библиотека. Вып. 1. Экология птиц: Виды, сообщества, взаимосвязи. Тр. науч. конф., посвящ. 150-летию со дня рождения Н.Н.Сомова (1861-1923). Харьков: 267-323.
• Волков С.В. 1997. Некоторые интересные орнитологические находки в верхнем течении Бикина // Рус. орнитол. журн. 6 (25): 7-11. EDN: KVWQFF
• Воробьёв К.А. 1954. Птицы Уссурийского края. М.: 1-360.
• Глущенко Ю.Н., Липатова Н.Н., Мартыненко А.Б. 2006а. Птицы города Уссурийска: фауна и динамика населения. Владивосток: 1-264.
• Глущенко Ю.Н., Нечаев В.А., Редькин Я.А. 2016. Птицы Приморского края: краткий фаунистический обзор. М.: 1-523.
• Глущенко Ю.Н., Шибнев Ю.Б., Волковская-Курдюкова Е.А. 2006б. Птицы // Позвоночные животные заповедника «Ханкайский» и Приханкайской низменности. Владивосток: 77-233.
• Джусупов Т.К. 2018. Оологические сборы Е.П. Спангенберга на юге Приморья, в центральной части, на севере и северо-востоке России // Selevinia 26: 107-129.
• Елсуков С.В. 1999. Птицы // Кадастр позвоночных животных Сихотэ-Алинского заповедника и Северного Приморья. Аннотированные списки видов. Владивосток: 29-74.
• Ильинский И.В. 1980. Сравнительно-экологический анализ приморских овсянок рода Emberiza в репродуктивный период. Автореф. дис. … канд. биол. наук. Л.: 1-20.
• Кулешова Л.В. 1969. Население птиц кедровых лесов Среднего Сихотэ-Алиня // Орнитология в СССР. Материалы 5-й Всесоюз. орнитол. конф. Ашхабад, 2: 344-347.
• Курдюков А.Б. 2014. Гнездовые орнитокомплексы основных местообитаний заповедника «Кедровая Падь» и его окрестностей: характер размещения и состояние популяций, дополнения к фауне птиц (материалы исследований 2008 года) // Рус. орнитол. журн. 23 (1060): 3203-3270. EDN: SWMORL
• Курдюков А.Б. 2017. Население птиц девственных неморальных хвойно-широколиственных лесов Южно-Уссурийского края: более полувека наблюдений // Динамка численности птиц в наземных ландшафтах. М.: 77-86.
• Кушнарёв Е.Л. 1984. Антропогенные сукцессии орнитосообществ и территориальные связи местообитаний западного Сихотэ-Алиня // Фаунистика и биология птиц юга Дальнего Востока. Владивосток: 71-78.
• Лабзюк В.И., Назаров Ю.Н., Нечаев В.А. (1971) 2020. Птицы островов северо-западной части залива Петра Великого // Рус. орнитол. журн. 29 (1981): 4626-4660. EDN: BXJMUK
• Лаптев А.А. 1984. Численность гнездящихся птиц в дубовых и долинных кедрово-широколиственных лесах Лазовского государственного заповедника // Исследования природного комплекса Лазовского заповедника. М.: 41-43.
• Литвиненко Н.М., Шибаев Ю.В. 1971. К орнитофауне Судзухинского заповедника и долины р. Судзухэ // Экология и фауна птиц юга Дальнего Востока. Владивосток: 127-186.
• Михайлов К.Е. 2014. Различия в заполнении тайги (сплошных массивов бореальных лесов) мелкими лесными птицами-мигрантами на примерах нескольких «модельных» для севера Приморского края групп видов Passeriformes. Часть 1 // Рус. орнитол. журн. 23 (978): 773-827. EDN: RXCEAD
• Михайлов К.Е., Шибнев Ю.Б., Коблик Е.А. 1998. Гнездящиеся птицы бассейна Бикина (аннотированный список видов) // Рус. орнитол. журн. 7 (46): 3-19. EDN: KTNORV
• Назаренко А.А. (1963) 2019. Зимняя орнитофауна юго-западного Приморья // Рус. орнитол. журн. 28 (1762): 1903-1912. EDN: QIDRBE
• Назаренко А.А. (1971) 2023. Краткий обзор птиц заповедника «Кедровая Падь» // Рус. орнитол. журн. 32 (2333): 3579-3631. EDN: QVHDNF
• Назаренко А.А. 1984. Птичье население смешанных и темнохвойных лесов Южного Приморья, 1962-1971 гг. // Фаунистика и биология птиц юга Дальнего Востока. Владивосток: 60-70.
• Назаренко А.А. 2014. Новое о гнездящихся птицах юго-западного Приморья: неопубликованные материалы прежних лет об орнитофауне Шуфанского (Борисовского) плато // Рус. орнитол. журн. 23 (1051): 2953-2972. EDN: QWKYLR
• Назаров Ю.Н. (2001) 2018. Распределение наземных гнездящихся птиц на островах Дальневосточного морского заповедника // Рус. орнитол. журн. 27 (1669): 4561-4569. EDN: UZEPVW
• Назаров Ю.Н. 2004. Птицы города Владивостока и его окрестностей. Владивосток: 1-276.
• Назаров Ю.Н., Казыханова М.Г., Катрич Л.Н. 1979. Летнее питание овсянок на юге Приморья // Биология птиц юга Дальнего Востока CCCР. Владивосток: 43-55.
• Назаров Ю.Н., Трухин А.М. (1985) 2020. К биологии сапсана Falco peregrinus и филина Bubo bubo на островах залива Петра Великого (Южное Приморье) // Рус. орнитол. журн. 29 (1987): 4884-4893. EDN: OWCJIG
• Назаров Ю.Н., Шибаев Ю.В. (1984) 2022. Список птиц Дальневосточного государственного морского заповедника // Рус. орнитол. журн. 31 (2212): 3329-3349. EDN: NODKXK
• Нечаев В.А. (2014) 2023. Птицы залива Восток Японского моря // Рус. орнитол. журн. 32 (2322): 3076-3099. EDN: XWCSUG
• Нечаев В.А., Курдюков А.Б., Харченко В.А. 2003. Птицы // Позвоночные животные Уссурийского государственного заповедника. Аннотированный список видов. Владивосток: 31-71.
• Нейфельдт И.А. 1970. Пуховые птенцы некоторых азиатских птиц // Тр. Зоол. ин-та АН СССР 47: 111-181.
• Омелько М.А. 1956. О перелётах птиц на полуострове Де-Фриза // Тр. ДВФ АН СССР 3, 6: 337-357.
• Панов Е.Н. 1973. Птицы Южного Приморья (фауна, биология и поведение). Новосибирск: 1-376.
• Пекло А.М. 2018. Птицы // Оологическая коллекция. Вып. 2. Воробьинообразные – Passeriformes. Черновцы: 1-224.
• Пукинский Ю.Б. (1974) 2011. Особенности гнездования овсянок, обитающих в бассейне реки Бикин // Рус. орнитол. журн. 20 (634): 366-367. EDN: NDBIAT
• Пукинский Ю.Б. 2003. Гнездовая жизнь птиц бассейна реки Бикин // Тр. С.-Петерб. общ-ва естествоиспыт. Сер. 4. 86: 1-267.
• Романов А.Л., Романова А.И. 1959. Птичье яйцо. М.: 1-620.
• Сотников В.Н. 2023. Каталог коллекций. Птицы – Aves. Оологическая и нидологическая коллекции. Вып. 2. Киров: 1-304.
• Спангенберг Е.П. 1940. Наблюдения над распространением и биологией птиц в низовьях реки Имана // Тр. Моск. зоопарка 1: 77-136.
• Спангенберг Е.П. (1965) 2014. Птицы бассейна реки Имана // Рус. орнитол. журн. 23 (1065): 3383-3473. EDN: SYCTWJ
• Тиунов И.М. 2004. Численность и распространение наземных гнездящихся птиц островов Римского-Корсакова // Дальневосточный морской биосферный заповедник. Биота. Т. 2. Гл. 5. Биота островов: распределение, состав и структура. Птицы островов Римского-Корсакова. Владивосток: 723-758.
• Харченко В.А. 2002. Зимнее население птиц Уссурийского заповедника и сопредельных территорий // Рус. орнитол. журн. 11 (186): 500-506. EDN: ISVQVP
• Харченко В.А. 2015. Население птиц основных лесных биотопов Южного Сихотэ-Алиня // Сиб. экол. журн. 4: 563-569.
• Шибнев Ю.Б. 1984. Фенологические наблюдения за птицами в заповеднике «Кедровая падь» // Фенологические явления в Приморье. Владивосток: 86-92.
• Шохрин В.П. 2008. Соколообразные (Falconiformes) и совообразные (Strigiformes) Южного Сихотэ-Алиня. Дис. … канд. биол. наук. Владивосток: 1-205 (рукопись).
• Шохрин В.П. 2014. Характеристика осеннего пролёта воробьинообразных в Лазовском заповеднике (юго-восток Приморского края) // Ареалы, миграции и другие перемещения диких животных. Владивосток: 372-381.
• Шохрин В.П. 2017. Птицы Лазовского заповедника и сопредельных территорий. Лазо: 1-648.
• Hoyt D.F. 1979. Practical methods of estimating volume and fresh weight of bird eggs // Auk 96: 73-77.
• Nartshuk E.P., Matyukhin A.V., Shokhrin V.P. 2022. Parasitic louse flies (Diptera, Hippoboscidae) and their association with bird hosts in the south of the Russian Far East // Entomol. Rev. 102, 3: 367-376.
• Nartshuk E.P., Matyukhin A.V., Shokhrin V.P. 2023. Birds as hosts of parasitic louse flies (Diptera) in the south of the Russian Far East // Зоол. журн. 102, 3: 310-316.
Юрий Николаевич Глущенко, Дмитрий Вячеславович Коробов. Тихоокеанский институт географии ДВО РАН, Владивосток, Россия. E-mail: yu.gluschenko@mail.ru; dv.korobov@mail.ru
Татьяна Владимировна Гамова. ФНЦ биоразнообразия наземной биоты Восточной Азии ДВО РАН, Владивосток, Россия. E-mail: birdsdv@mail.ru
Валерий Павлинович Шохрин. Объединённая дирекция Лазовского государственного природного заповедника им. Л.Г. Капланова и национального парка «Зов тигра», с. Лазо, Приморский край, Россия. E-mail: shokhrin@mail.ru
Николай Николаевич Балацкий. Новосибирск, Россия. E-mail: nnbal54@mail.ru
Геннадий Николаевич Бачурин. Научно-практический центр биоразнообразия, Ирбит, Свердловская область, Россия. E-mail: ur.bagenik@mail.ru
Владимир Несторович Сотников. Кировский городской зоологический музей, Киров, Россия. E-mail: sotnikovkgzm@gmail.com
Дмитрий Анатольевич Беляев. Приморский государственный аграрно-технологический университет, Уссурийск, Приморский край, Россия. Объединённая дирекция государственного природного биосферного заповедника «Кедровая падь» и национального парка «Земля леопарда» им. Н.Н. Воронцова, Владивосток, Россия. E-mail: d_belyaev@mail.ru
Иван Михайлович Тиунов. ФНЦ биоразнообразия наземной биоты Восточной Азии ДВО РАН,
Владивосток, Россия. Государственный природный биосферный заповедник «Ханкайский», Спасск-Дальний, Приморский край, Россия. E-mail: ovsianka11@yandex.ru
Источник
Русский орнитологический журнал 2024, экспресс-выпуск 2425. стр. 2495-2525.