

Литературные сведения о биологии теньковки Phylloscopus collybita в лесостепи Западной Сибири до сих пор фрагментарны (Пукинский 1969; Гынгазов, Миловидов 1977). Подробно проанализированы лишь особенности миграций этого вида (Чернышов 2004). Материал собран в 1973-2003 годах в Здвинском районе Новосибирской области, на территории, прилегающей к юго-восточному побережью озера Малые Чаны. Сезонные перемещения пеночки-теньковки изучали путём отловов птиц стационарными линиями «паутинных» сетей (Юрлов и др. 1975) и методом прямых визуальных наблюдений с постоянного наблюдательного пункта (НП) (Гаврилов 1977).
В период гнездования обследовано и находилось под наблюдением 24 гнезда, промерено 92 яйца. Для оценки изменчивости яиц определялись с помощью штангенциркуля (точность деления 0.1 мм) их линейные размеры: длина (L) и максимальный диаметр (B). Описание состояния оперения во время линьки проводили по методике Г.А. Носкова с соавторами (Носков, Гагинская 1972; Носков, Рымкевич 1977) по несколько сокращённой схеме (не описывалось состояние оперения на некоторых мелких участках птерилий). Всего сделано 20 описаний линьки взрослых и 63 – молодых особей. При обработке данных по полной линьке взрослых птиц выделяли 11 стадий (десять по началу замены каждого первостепенного махового пера, а последняя, 11-я стадия – этап дорастания оперения после отрастания самого дистального махового).
В районе исследований теньковка – обычный гнездящийся и многочисленный пролётный вид. Прилетает во второй-третьей декаде апреля. Самая ранняя дата прилёта – 13 апреля. Наиболее интенсивно теньковки летят в первой половине мая с «пиком» пролёта во второй пятидневке. Во второй половине мая интенсивность пролёта постепенно снижается, а в первой декаде июня миграция этого вида практически заканчивается (рис. 1).
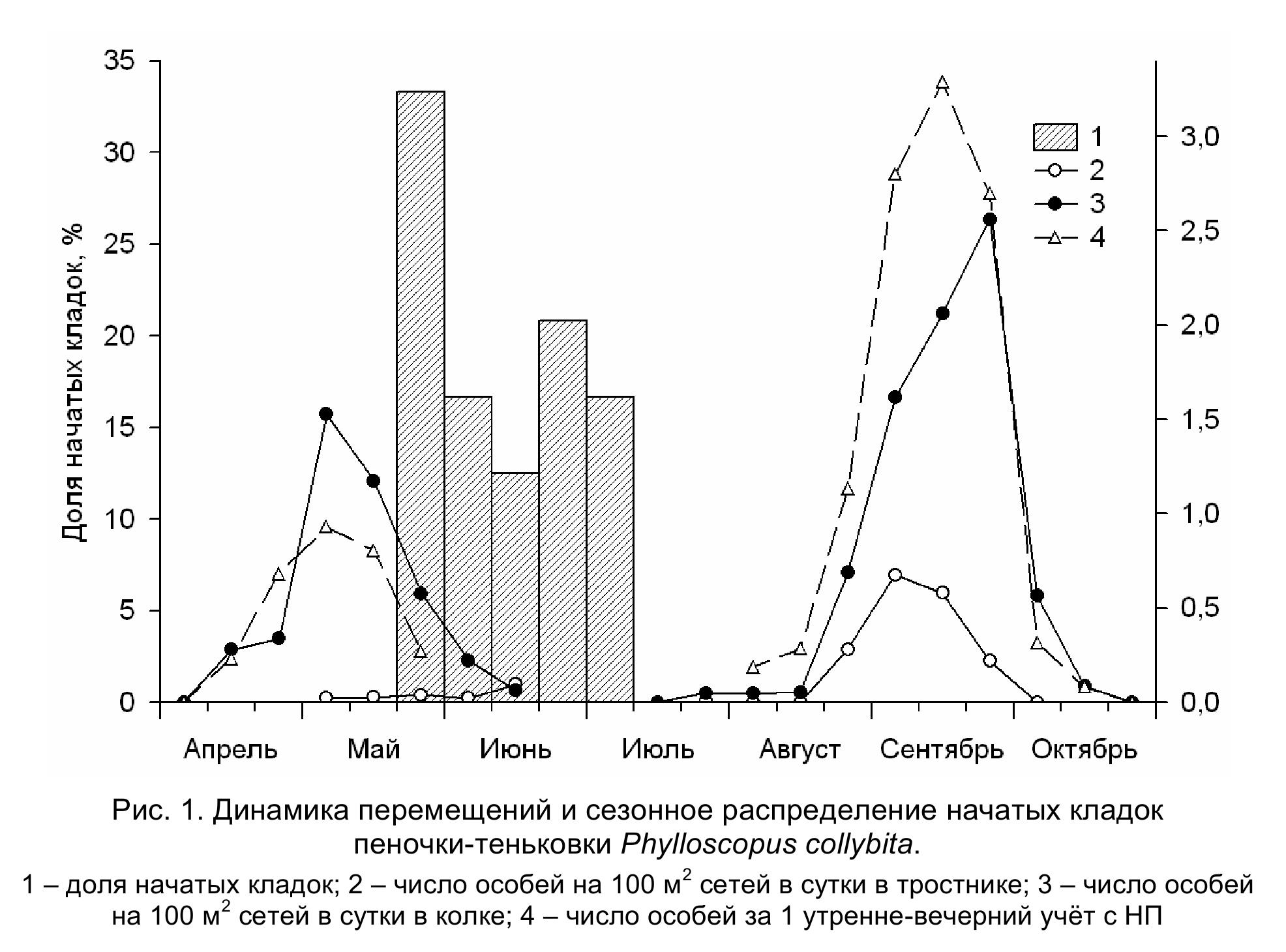
Теньковка населяет различные типы берёзовых и осиново-берёзовых перелесков-колков, нередко заболоченных, предпочитая более разреженные, с густым подлеском из кустов ивы, чёрной смородины и шиповника. Из 24 найденных гнёзд теньковки лишь одно располагалось на земле в зарослях крапивы, остальные – на высоте до 56 см. 12 гнёзд (50 %) были построены на шиповнике, 5 (20.8 %) – на чёрной смородине, по 2 – на стеблях крапивы и высоких злаков, единичные гнёзда найдены на ветках ивы и небольшой осинки. Средняя высота размещения гнёзд – 27±4 см. Внешние размеры гнезда, имеющего форму шара с боковым летком, следующие, мм: «диаметр» от 85 х 80 до 130 х 105 и 117 х 117, в среднем 107±3 х 96±3; высота 90-135, в среднем 102±3; размеры летка от 34 х 27 до 50 х 38, в среднем 41±2 х 33±1. Гнездо строится из сухих травинок, лоток обильно выстилается перьями. Параметры гнёзд соответствуют значениям, приводимым в литературе (Schцnfeld 1978; Лапшин 2000). Несмотря на ранний прилёт, к гнездованию теньковки приступают относительно поздно. Период откладки яиц у теньковки растянут на 1.5 месяца (рис. 1). В самом раннем гнезде первое яйцо отложено 25 мая, в самом позднем – 9 июля.
В полной кладке теньковки 3–6, в среднем 5.10±0.19 яиц. Чаще всего встречаются кладки с 5 и 6 яйцами (52.4 % и 33.3 % соответственно). Величина кладки теньковки в окрестностях озера Чаны, вероятно, меньше, чем в большинстве других частей её ареала, за исключением Ленинградской области (Schцnfeld 1978; Гуреев, Миловидов 1983; Лапшин 1983, 2000; Мальчевский, Пукинский 1983; Зимин 1988; Гуреев 1989). При этом в течение сезона наблюдается существенное уменьшение средней величины кладки: от 5.71±0.49 яиц в последней декаде мая до 4.25±0.48 яиц, отложенных в первой декаде июля (P < 0.01).
Яйца белые с красновато-бурыми крапинами. Размеры яиц, мм: от 13.3 х 11.2 и 14.7 х 11.1 до 16.5 х 11.6 и 15.9 х 13.0, в среднем 15.51±0.06 х 11.88±0.04. Вероятно, сходные размеры и форма яиц у теньковок в Карелии и на Западном Алтае (Лапшин 2000; Щербаков 2009), но в Германии теньковки откладывают более округлые яйца, т.е. в среднем меньшей длины, но большего диаметра (Schцnfeld 1978). Коэффициент вариации (CV) длины равен 3.5 %, наибольшего диаметра (ширины) – 3.0 %. Индекс округлённости составляет 70.3-84.2 %, в среднем 76.6±0.3 %, CV = 3.7 %. Объём яиц изменяется в пределах от 851 до 1379 мм3, в среднем равняясь 1118±9 мм3 при CV = 8.1 %. Каких-либо существенных сезонных изменений в размерах и форме откладываемых яиц у теньковки не обнаружено. Из 17 гнёзд теньковки с известной судьбой гнездовой цикл успешно завершён только в 6 (35.3 %); 9 гнёзд (52.9 %) разорено хищниками, в одном все 6 яиц оказались неоплодотворёнными, а в другом вылупившиеся птенцы по неизвестной причине погибли. Выводится 2-6, в среднем 4.4±0.3 птенцов, а покидают гнездо 1-6, в среднем 3.5±0.8 слётков. Доля неразвившихся яиц (неоплодотворённых и с погибшими на разных стадиях развития эмбрионами) от общего количества яиц в кладках, сохранившихся по крайней мере до вылупления, у теньковки составляет 19.7±5.6%, а без учёта гнездб, где все яйца были «болтунами» – 12.9±4.0 %. Эмбриональная элиминация отмечена в 9 из 15 кладок. Гибель 1-4 птенцов (постнатальная смертность) зарегистрирована в 3 из 7 выводков. Успешность размножения (доля вылетевших птенцов от числа отложенных яиц) у теньковки составляет лишь 28.8 %.
Сезон полной послебрачной линьки у теньковки растянут с первой декады июля до конца второй декады сентября. У пойманной 10 июля самки зарегистрирована вторая стадия линьки. Ещё у двух самок, отловленных 16 и 23 июля, линька была на четвёртой стадии. С 26 августа по 17 сентября отлавливались теньковки на 11-й стадии линьки, хотя полностью перелинявшие взрослые особи встречаются уже с третьей декады августа.
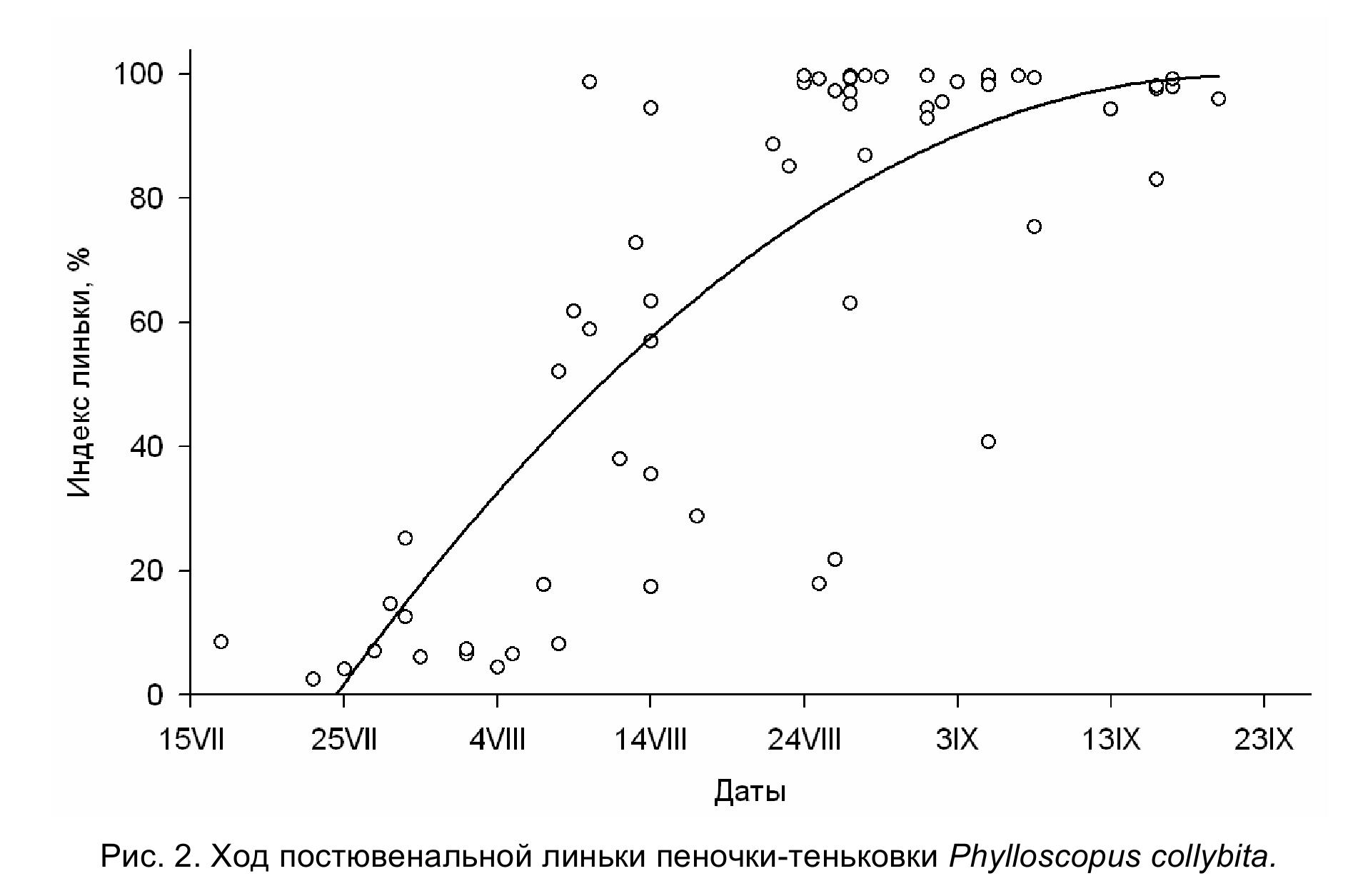
Молодые теньковки приступают к смене юношеского оперения во второй декаде июля. Наблюдается довольно сильный «разброс» в индивидуальных сроках начала линьки (рис. 2), вероятно обусловленный растянутостью сезона гнездования. Во время линьки заменяются контурные перья на туловище и голове, перья на плечевой, бедренной и голенной птерилиях, верхние и нижние кроющие хвоста, зарастают пуховидным пером аптерии. Среди верхних кроющих крыла сменяются средние и малые верхние кроющие второстепенных маховых, кроющие пропотагиума. У части особей линяют кроющие перья третьестепенных маховых, но сами эти маховые не заменяются. Из нижних кроющих крыла заменяются нижние маргинальные кроющие и средние нижние кроющие второстепенных маховых. У некоторых птиц линяет центральная пара рулевых перьев. Незакончившие линьку особи отлавливались до начала третьей декады сентября, а уже перелинявшие – начиная с третьей декады августа. В послегнездовой период первые кочующие теньковки попадались в сети во второй половине июля. Последние пролётные птицы отловлены в середине октября (самая поздняя дата – 14 октября). Период интенсивных перемещений значительно короче и длится с пятой пятидневки августа по первую пятидневку октября. Во время пролёта пеночки-теньковки в значительном количестве встречаются в тростниковых займищах и в четырёхрядных лесополосах из тополя.
Автор благодарен К.Т. Юрлову и В.М. Тотунову, оказавшим помощь в полевых исследованиях.
Литература
• Гаврилов Э.И. 1977. Методика сбора и обработки материалов по количественной характеристике видимых миграций птиц // Методы изучения миграций птиц. Материалы Всесоюзной школы-семинара. М: 96-117.
• Гуреев С.П. 1989. Величина кладки и успешность размножения птиц в Кузнецком Алатау // Биопродуктивность и биоценотические связи наземных позвоночных юго-востока Западной Сибири. Томск: 56-74.
• Гуреев С.П., Миловидов С.П. 1983. Материалы по экологии славковых (Sylviidae) в Западной Сибири // Экология наземных позвоночных Сибири. Томск: 105-119.
• Гынгазов А.М., Миловидов С.П. 1977. Орнитофауна Западно-Сибирской равнины. Томск: 1-350.
• Зимин В.Б. 1988. Экология воробьиных птиц Северо-Запада СССР. Л.: 1-184.
• Лапшин Н.В. (1983) 2008. Материалы по размножению четырёх видов пеночек Phylloscopus на севере Ленинградской области и в южной Карелии // Рус. орнитол. журн. 17 (398): 156-157.
• Лапшин Н.В. 2000. Биология теньковки Phylloscopus collybita в Карелии // Рус. орнитол. журн. 9 (90): 3-27.
• Мальчевский А.С., Пукинский Ю.Б. 1983. Птицы Ленинградской области и сопредельных территорий: история, биология, охрана. Л., 2: 1-504.
• Мянд Р. 1988. Внутрипопуляционная изменчивость птичьих яиц. Таллин: 1-192.
• Носков Г.А., Гагинская А.Р. 1972. К методике описания состояния линьки у птиц // Сообщ. Прибалт. комис. по изучению миграций птиц 7: 154-163.
• Носков Г.А., Рымкевич Т.А. 1977. Методика изучения внутривидовой изменчивости линьки у птиц // Методики исследования продуктивности и структуры видов птиц в пределах их ареалов. Вильнюс, 1: 37-48.
• Пукинский Ю.Б. 1969. Воробьиные птицы естественных ландшафтов Барабинской низменности // Вопросы экологии и биоценологии 9: 62-78.
• Чернышов В.М. 2004. Миграции пеночек в Барабинской лесостепи // Миграции птиц в Азии. Алматы, 12: 166-184.
• Щербаков Б.В. 2009. К экологии теньковки Phylloscopus collybita на Западном Алтае // Рус. орнитол. журн. 18 (527): 2043-2047.
• Юрлов К.Т., Тотунов В.М., Чернышов В.М. 1975. Опыт отлова птиц «кустарниковыми» и «кронными» сетями в Барабинской лесостепи (Западная Сибирь) // Материалы Всесоюз. конф. по миграциям птиц. М., 2: 131-132.
• Hoyt D.F. 1979. Practical methods of estimating volume and fresh weight of bird eggs // Auk 96, 1: 73-77.
• Rymkevich T.A., Bojarinova J.G. 1996. Variation in the extent of postjuvenile moult in the Great Tit near Lake Ladoga (Russia) // Bird Study 43: 47-59.
• Schцnfeld M. 1978. Der Weidenlaubsдnger Phylloscopus collybita // Die Neue BrehmBьcherei 511. Wittenberg Lutherstadt: 1-136.
Источник
Русский орнитологический журнал 2011, Экспресс-выпуск 673. стр. 1419-1424.