

Семейство: Фазановые Phasianidae
Род: Фазаны Phasianus
Вид: Phasianus colchicus (Linnaeus, C 1758)
Протоним: Phasianus colchicus.
Синоним: Phasianus colchicus, Phasianus colchicus [colchicus Group], Phasianus colchicus [colcichus Group], Phasianus colchicus colchicus.
Немецкое название: Jagdfasan
Испанское название: Faisán vulgar.
Казахское название: кәдімгі қырғауыл, Қырғауыл
Украинское название: фазан звичайний.
Другие названия: обыкновенный фазан, охотничий фазан.
Taxonomic Serial Number : 175905 | Global Biodiversity Information Facility : 9752149.
Список ПОДВИДОВ вида Фазан Phasianus colchicus (Linnaeus, C 1758)
‐ Северокавказский фазан — Phasianus colchicus septentrionalis (Lorenz, T 1889)
‐ Закавказский фазан — Phasianus colchicus colchicus (Linnaeus, C 1758)
‐ Талышский фазан — Phasianus colchicus talischensis (Lorenz, T 1889)
‐ Персидский фазан — Phasianus colchicus persicus (Severtsov, NA 1875)
‐ Мургабский фазан — Phasianus colchicus principalis (Sclater, PL 1885)
‐ Хивинский фазан — Phasianus colchicus chrysomelas (Severtsov, NA 1875)
‐ Аму-Дарьинский фазан — Phasianus colchicus zarudnyi (Buturlin, SA 1904)
‐ Таджикистанский фазан — Phasianus colchicus bianchii (Buturlin, SA 1904)
‐ Зеравшанский фазан — Phasianus colchicus zerafschanicus (Tarnovski, GV 1893)
‐ Сыр-Дарьинский фазан — Phasianus colchicus turcestanicus (Lorenz, T 1896)
‐ Семиреченский фазан — Phasianus colchicus mongolicus (von Brandt, JF 1844)
‐ Маньчжурский фазан — Phasianus colchicus pallasi (Rothschild, LW 1903)
ОСНОВНОЕ ОПИСАНИЕ ПО: Фазан — Phasianus colchicus (Linnaeus, C 1758)
Окраска. Взрослый самец (номинативный подвид). Общая окраска красновато-золотистая, крылья светло-коричневые, голова зеленая с металлическим отливом, передняя часть шеи и верх груди фиолетовые, с таким же отливом. На удлиненных золотисто-коричневых перьях затылка узкие вершинные зеленые каймы. Зашеек фиолетовосиний, с зеленоватым металлическим блеском. На красновато-золотистом фоне верхней части тела (с фиолетовым отливом на пояснице и верхних кроющих крыла) расположен сложный узор из черных пятен и чешуевидного рисунка в передней части спины, а также белых с черным окаймлением стрельчатых пятен в лопаточной области. Все перья верхней части тела несут широкие красноватые каймы, которые под разными углами зрения выглядят то фиолетовыми, то золотистыми. Эти каймы очень широки на перьях поясницы и надхвостья и образуют здесь однородную золотисто-рыжую поверхность с тем же фиолетовым отливом. Нижняя часть тела более светлая, золотистая, с густым чешуйчатым рисунком из черных полукружий на груди и из поперечных черных полос и пятен на передней части брюха и боках. Брюхо черно-бурое. Маховые перья светло-коричневые, с нечетким поперечным рисунком из беловатых полос. Плечевые и кроющие перья крыла темно-коричневые, с белым поперечнополосатым узором и широкими желтовато-серыми краями, имеющими узкий коричневый или фиолетовый (в зависимости от угла зрения) бордюр.
Рулевые перья желтовато-коричневые с узкими охристо-рыжими или фиолетовыми каймами на 3-х центральных парах и с поперечным рисунком из узких черных полос, которые в нижней части центральной пары не доходят до краев. На крайних парах полосы эти более широкие, и к ним добавляется коричневый струйчатый рисунок. Клюв и ноги желтоватые, на боках головы участки голой кожи, лишенные перьев, захватывающие область вниз от глаза до ушных отверстий и клюва — «щеки», становящиеся ярко красными в период токования.
Взрослая самка. Окраска пестрая, «рябая». На верхней части тела по песочному фону в правильном порядке расположены овальные черновато-коричневые пятна — центры перьев. На голове и зашейке пятна принимают вид поперечных полосок и расположены тесно друг к другу, отчего эти части выглядят более темными. На перьях головы и зашейка имеется слабый фиолетовый блеск. Нижняя часть тела светло-песочного цвета со слабым струйчатым рисунком на брюхе и с полукруглыми бурыми пятнами в верхней части груди и нижней части шеи. На боках тела тот же рисунок из продолговатых овальных пятен бурого цвета по серо-желтому фону. Брюхо одноцветное, с едва заметным желтоватым струйчатым рисунком. Маховые перья окрашены, как и у самцов, но с более четким поперечным рисунком из белых полос. Кроющие перья крыла окрашены так же, как и на спине. Рулевые перья с четким поперечным рисунком из тонких белесых и окаймляющих их более широких черно-бурых полос, которые на центральной паре не доходят до краев — здесь идет широкое серое окаймление с тонкой коричневой струйчатостью. Клюв и ноги серые, голые пятна по бокам головы небольшие, идут от глаз до ушных отверстий.
Самец и самка в ювенальном наряде. Окрашены одинаково. Общий тон наряда, как и у взрослой самки, песочно-серый с черно-бурыми и коричневыми пестринами. Верх головы и зашеек в мелких коричнево-бурых пестринах, на передней части спины — крупные поперечные и продольные полосы темно-коричневого цвета с продольной белой штриховкой, создаваемой узкими наствольными полосками. В лопаточной области на темно-буром фоне центральной части перьев их широкие светлые каймы создают ланцетовидный рисунок. На пояснице и надхвостье неясные буроватые поперечные полосы. Нижняя часть тела песочного цвета с поперечным рисунком из коричневых полос на боках и небольшим количеством пестрин в верхней части груди. Горло беловатое. Маховые и рулевые перья с поперечным рисунком из светлых полос по буроватому фону, и только внутренние второстепенные маховые и плечевые перья несут сложный ланцетовидный рисунок в верхней части пера.
Второй ювенальный наряд. Формируется партиями перьев, вырастающих в последнюю очередь. Он называется некоторыми авторами «переходным», но в действительности отличается от наряда взрослого не менее, чем и первый ювенальный. Его формируют перья на груди, брюхе, на зашейке и спине: они имеют ярко-рыжие каймы и бледно-охристые пятна вдоль черного настволья, причем между каймами и центром пера идет черная полоска. Эти перья создают преобладающую рыжую окраску груди и брюха. Сразу же с появлением этого наряда начинается интенсивный рост взрослого оперения (Тугаринов, Козлова, 1945).
Пуховые птенцы снизу светло-серые, с легким желтоватым оттенком и с продольным коричневым рисунком из широких полос по верхней части тела: центральная коричневая полоса идет от темени, заостряясь спереди назад, по середине спины и поясницы до хвостовой области. По бокам ее тянутся 2 светлые полосы, окаймленные снаружи еще парой темно-бурых полос, начинающихся в надорбитальной области. По бокам головы темно-бурые пятна в области уха.
Строение и размеры. Основные особенности строения фазана — короткие, сильно закругленные крылья, сильные ноги значительной длины и длинный ступенчатый хвост, где самой длинной является центральная пара рулевых перьев, причем все они имеют заостренные тонкие вершины.
По бокам головы над ушными перьями у самцов расположены 2 пучка удлиненных перьев, образующих так называемые «уши». Клюв крепкий, сильный, слегка загнут вниз.
Самцы (п = 29, кол. ЗИН АН СССР):
крыло 253.5 (228—263),
хвост 543 (457—628),
длина клюва 21.0 (18.1—22.4),
плюсна 71.5 (61.7—75.2),
средний палец 49.5 (45.7—52.4).
Самки (п=10, кол. ЗИН АН СССР):
крыло 226.1 (214—236),
хвост 335.5 (283—433),
длина клюва 17.9 (16.6—19.4),
плюсна 63.1 (59.7—68.1),
средний палец 45.5 (40.5—50.0).
Масса самцов и самок варьирует в зависимости от возраста, подвидовой принадлежности и сезона — у самцов от 900 до 2000, у самок от 710 до 1350.
Суточная активность, социальное поведение. Суточный ритм активности практически не меняется весь год и представляет собой чередование периодов повышенной кормовой активности утром и вечером с дневным отдыхом и ночным сном. Фазаны — стайные птицы и, за исключением периода размножения, держатся небольшими разреженными группами, образуя в конце лета и осенью довольно большие скопления на особо благоприятных для кормежки местах. В заповеднике «Тигровая балка» например, в сентябре—октябре на отдельных участках сенокосов среди тростниковых зарослей собиралось по нескольку десятков птиц. О ночевках зимой говорилось выше. Птицы кавказских популяций летом ночуют на деревьях.
Хозяйственное значение, охрана. Существованию фазана как виду ничто не угрожает благодаря практике широкого разведения так называемого охотничьего фазана. В то же время судьба аборигенных форм, в том числе всех, обитающих в нашей стране, вызывает большую тревогу.
Основные причины, угрожающие их существованию — химизация сельского хозяйства, бесконтрольная охота и поглотительное скрещивание с «охотничьим» фазаном, выпускаемым деятелями охотничьих хозяйств в условиях полной анархии и отсутствия научного контроля. Уничтожение исконных мест обитания фазанов — тугайной растительности, не столь уж трагично, поскольку фазан легко уживается с человеком при отсутствии преследования с его стороны, и находит все для себя необходимое в агроценозах, особенно там, где сады чередуются с полями и небольшими участками дикорастущих кустарников. Высокая оседлость фазанов делает их перспективными для охотничьих хозяйств, а эффективность самых простых биотехнических мер (посев ягодных кустарников, подкормка зимой, охрана гнездовых территорий) сразу же дает хороший эффект.
До тех пор, пока существует угроза со стороны различных химикатов, необходимо для каждого подвида, пока не поздно, создать питомники по вольерному разведению, что легко выполнимо на базе существующих заповедников и заказников.
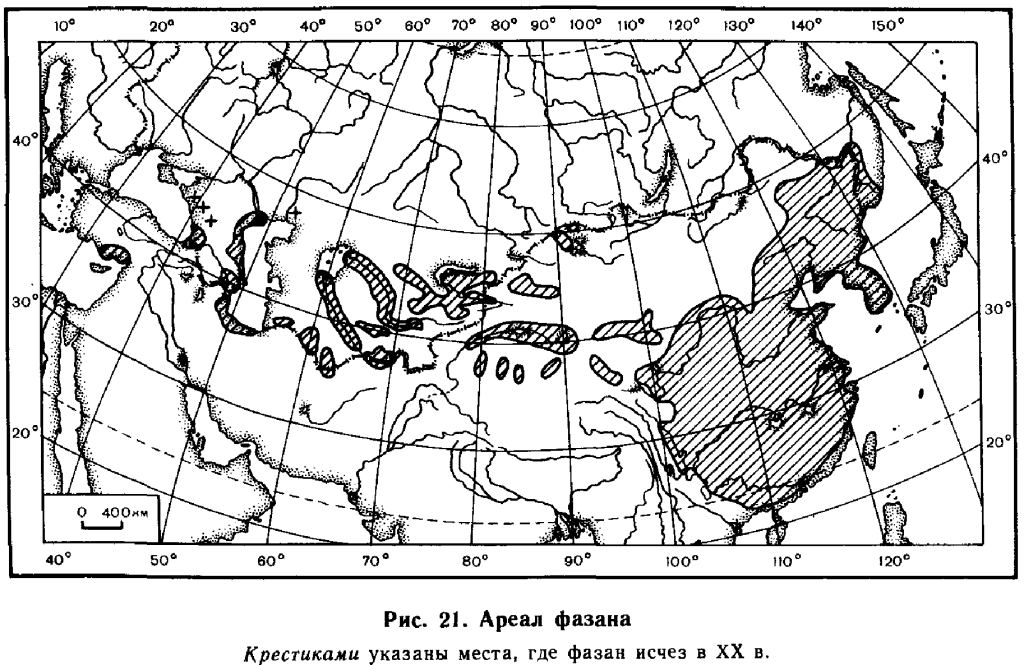

В древности фазан был завезен в Грецию и распространился оттуда на Балканы. Впоследствие с помощью человека фазаны были широко расселены по всей Европе, за исключением Фенноскандии, в Северной Америке, на Гаваях, в Новой Зеландии. Они были также успешно интродуцированы кое-где в Южной Америке (Чили), в Южной Австралии и Тасмании (Long, 1981). В СССР первоначальное распространение охватывало Предкавказье и Закавказье, прибрежные районы Каспийского моря, тугайные заросли в ряде мест Казахстана и Средней Азии, а также долины ряда рек на юге Дальнего Востока, к северу до долины Амура включительно.
Сейчас, как уже указывалось выше, искусственное разведение гибридного, так называемого охотничьего фазана, привело к его расселению во многих областях юга Европейской части СССР, а также кое-где в Средней Азии.
Численность. В настоящее время почти повсеместно аборигенные формы фазана находятся на грани уничтожения и имеют очень низкую численность. Только искусственно разводимый охотничий фазан, успешно размножаясь в специальных угодьях, бывает довольно многочисленным.
Основной причиной низкой численности фазана является непрекращающееся преследование со стороны человека и, в меньшей степени, уничтожение естественных мест его гнездования — тугайных зарослей. В благоприятных же местах, при отсутствии названных причин, фазаны достигают очень высокой численности и способны существовать при высокой плотности населения. Например, в 1940-х гг. в заповеднике «Тигровая балка» число выводков на 1 км2 тугаев достигало 20, а число птиц здесь же осенью — 140—150 особей на 1 км2 (Тугаринов, Козлова, 1945). Примерно такую же картину я застал в этом заповеднике в 1958—1959 гг.: 15 территориальных самцов на 100 га весной и до 120 птиц на ту же площадь осенью. Сейчас в том же заповеднике в наиболее благоприятных местах плотность составляет весной 2—6 самцов на 100 га (Бидос, 1985).
В большинстве же районов Средней Азии, Казахстана и Кавказа численность их в настоящее время на 100 га не превышает 1—3 территориальных самцов весной и 5—15 птиц различного пола и возраста осенью. Еще в сравнительно недавнем прошлом этим птицам были свойственны периодические подъемы численности, нередко приводившие к расширению или восстановлению ареала в отдельных его частях, вслед за которыми могли наступать и новые депрессии. Однако естественный ход динамики численности был полностью нарушен человеком еще до того, как его успели изучить. Поэтому далеко не выяснены факторы, влияющие на численность фазана в естественных условиях, в том числе и те, которые вызывают ее резкие подъемы или спады. Сейчас можно только сказать, что основными причинами, отрицательно влияющими на численность птиц, помимо антропогенных факторов являются стихийные бедствия разного рода, особенно пожары и наводнения в период инкубации яиц, а также деятельность различных хищников (прежде всего млекопитающих).
В поисках корма птицы роются в подстилке, разгребая ее ногами, но корни и клубни в рационе почти не представлены. Только при отсутствии ягод (например, в дельте Волги) луковицы, клубеньки камыша и стрелолиста, корни щавеля и жерухи поедаются в большом количестве (Шатас, 1963). В Приморье фазаны выкапывают на полях мелкий картофель (Гладков, 1952).
Сезонная смена кормов выражена нерезко, но можно отметить значительное преобладание в зимнем рационе различных ягод (особенно в начале зимы) и семян различных растений, в основном бобовых, гречишных и злаков, в том числе культурных. Среди ягод и плодов особенно важны джида, облепиха, солянки, ежевика, шиповник, дереза, саксаул, джузгун, боярышник, эфедра, барбарис. Местами (Приморье) важное место занимают желуди. Как исключение наблюдалось также использование в суровые многоснежные зимы почек ивы, туранги и сережек березы (Или, Уссурийский край). В различных районах зимняя диета бывает несколько различной, будучи обусловлена флористическими особенностями той или иной местности. По долине Сырдарьи например, основные зимние корма — это ягоды джиды и семена чингила, шерстистой солянки и петросимонии (Тюреходжаев, 1974), в долине Или — ягоды джиды и облепихи и семена маревых (кохии, сведы, терескена, соляиок), бобовых (чингила, солодки, люцерны, донника) и злаков (тимофеевки, пырея и т.п.). На Иссык-Куле главный зимний корм — ягоды облепихи, в тугаях верховьев Амударьи — ягоды джиды, в Центральном Тянь-Шаие (верховья р. Текес) — ягоды облепихи, барбариса, шиповника, листья и семена восточного ломоноса, различные семена (Жирнов, Винокуров, 1959). Весной и летом на Зеравшане особенно важны семена Centaurea (Мекленбурцев, 1940). Наряду с большим количеством различных семян поедаются также ягоды тута, самая различная зелень — листья солодки, бобовых и, наконец, большое количество разнообразных насекомых. На первом месте среди них стоят саранчовые, истребляя которых фазаи несомненно приносит большую пользу. Из других насекомых важное место занимают муравьи, различные мелкие жуки, нимфы цикад, палочники, а также пауки. В Уссурийском крае птицы весной часто питаются прорастающими желудями манчжурского дуба: находили до 30 желудей в одном зобу (Слепцов, Горчаковская, 1952). На Кавказе в июне важное значение имеют моллюски, в августе очень важны семена ежевики, лиции, лоха, к которым добавляются насекомые, в основном саранчовые. В дельте Волги насекомые составляют 70 % объема суточного рациона, причем преобладают долгоносики, златки, жужелицы, кузнечики и муравьи. На Сырдарье летом животные корма составляют около 60 %, а в горных районах ягоды облепихи становятся главным кормом уже летом. Осенью в рационе преобладают различные ягоды, всевозможные семена и плоды. Растительные корма составляют в этот сезон 80 % рациона (Сырдарья) и больше.
Примечательно, что во многих местах насекомые встречаются в пище фазана и зимой.
Птенцы в первые дни питаются исключительно насекомыми. Они проявляют большую избирательность, поедая преимущественно личинок мелких саранчовых, тлей, муравьев, пауков. Насекомых с жестким хитиновым покровом поедают только с 7-го дня жизни (Тугаринов, Козлова, 1945).
Наиболее типичны участки, где имеются сочетания густых кустарниково-травянистых зарослей, открытых полян и водопоев. В зарослях устраивается гнездо, и там же птицы ночуют, а на открытых местах с невысокой травой они кормятся. Особенно благоприятны для гнездования окраины густых тростниково-кустарниковых тугаев вблизи водопоев, граничащие с открытыми местами — пойменными лугами или полупустыней.
Если ближайшие водоемы находятся на некотором удалении от гнездовых участков, то каждая пара регулярно ходит на водопой по определенным маршрутам. На гнездовом участке у самца имеется 2—3 точки, на которых он токует особенно часто и откуда ведет наблюдение за соседями. В пределах своего участка самец все время перемещается, причем по вполне определенному маршруту. В «Тигровой балке» такой маршрут составлял в среднем 400—500 м, и самец проходил его за 30—40 мин, как бы непрерывно совершая обход своего участка (Абдусалямов, 1971). Самка располагает гнездо в самом глухом уголке участка. Весеннее оживление начинается рано: в феврале — в южном Таджикистане, в начале марта — на Балхаше. Однако созревание гонад происходит значительно позже, в конце марта—начале апреля. В это время брачная активность самцов максимальна. Крики самцов звучат особенно часто на восходе, днем они стихают, а вечером вновь становятся более частыми. Интервалы между токовыми криками в это время равняются в среднем 6 мин, позже увеличиваются до 10 мин (Тугаринов, Козлова, 1945). В это время часто происходят стычки между самцами, обычно между владельцами участка и посторонним. Стычки могут принимать довольно ожесточенный характер, и, как правило, заканчиваются победой хозяина. В начале схватки самцы стоят друг против друга, вытянув навстречу головы и шеи, клюв к клюву, совсем как домашние петухи. При драке удары наносятся крыльями, клювами и ногами, но прежде всего каждый из соперников норовит подпрыгнуть повыше, чтобы нанести удар ногами сверху. Есть непроверенные сообщения о том, что некоторые самцы погибают от ран, полученных в таких схватках. Драке нередко предшествует особая конфронтация, когда самцы бегут бок о бок некоторое расстояние (от 5 до 15 м), так называемая параллельная дуэль. Активность токования быстро гаснет к началу мая и полностью затухает к концу мая, к моменту вылупления птенцов. С этого времени территориализм исчезает полностью, самцы покидают гнездовые участки и перемещаются в другие места, в густые заросли вблизи водоемов, где у них протекает линька. Осенее токование выражено слабо, брачные крики можно снова слышать в сентябре—октябре.
Брачный ритуал самца довольно прост (рис. 23).


Громкий брачный крик, слышимый за 400—600 м, представляет собой двусложный, несколько картавый и в то же время звонкий звук, где последний слог, а иногда первый, состоит как бы из двух слившихся слогов, так что в целом крик этот звучит как «кх. . . кх-кх». Почти синхронно с этими слогами или сразу вслед за ними (у разных подвидов, видимо, существуют определенные различия в этом дисплее) следует короткая, около 0.5 с серия активных взмахов крыльями, хлопки которых сливаются в вибрирующий звук, вроде «прррр». Крылья работают так быстро, что отдельных взмахов различить не удается, причем во время взмахов передняя часть тела птицы слегка поднимается, и бьющие крылья не касаются земли. В этот момент хвост приподнимается на 45—50°, рулевые перья раскрываются, но немного, продолжая краями налегать друг на друга. Другой вокальный элемент тока — негромкое «гуканье», звучащее как довольно глухое «гу-гу». Этот звук выражает особенно сильное возбуждение и обычно издается самцом в непосредственной близости от самки. Находясь перед самкой, самец начинает рыхлить землю клювом, делает клевки, подбирая какое-нибудь семечко и вновь бросая перед собой, как бы приглашая самку покормиться. После такого привлечения самки и ее кормления нередко происходит копуляция. Ей предшествует резкое набухание и покраснение голых участков кожи на голове самца, а пучки перьев — «уши» расходятся и торчат в стороны. После копуляции самец взъерошивает оперение, слегка приподнимает одно крыло и опускает другое, так что оба крыла и спина образуют одну плоскость, наклоненную в сторону самки.
Хвост раскрывается веером, приподнимается и тоже поворачивается в той же плоскости, что и крылья, шея сильно вздувается и изгибается дугой, а голова наклоняется так, что клюв почти касается земли. В такой позе самец медленными шагами делает вокруг самки полукруг, непрерывно издавая громкое шипение и производя особый шум быстрыми колебаниями рулевых перьев (Тугаринов, Козлова, 1945).
Гнездо располагается в глухих зарослях и обычно хорошо скрыто сверху густым переплетением трав, формирующим своеобразную крышу. Оно имеет обычно два входа, и самка при нападении хищника всегда может спастись, используя второй вход. Лоток бывает довольно глубоким и хорошо выстлан сухими травами. Диаметр лотка 20—23 см, глубина его 5—7 см (Строганов, 1946). На Дальнем Востоке и в Японии гнездо нередко выстилается сухими листьями. Приверженность самки к избранному ею участку довольно сильная. Нередки случаи, когда одна и та же гнездовая ямка используется несколько лет подряд, и такие гнезда насчитывают соответствующее число слоев с остатками скорлупы — каждый раз новая выстилка делается поверх старой (Видос, 1985). Количество яиц в кладке колеблется от 8 до 24; на Сырдарье, например, в 20-х гг. средний размер кладки составлял 20 яиц (Горчаковский, 1928). Повторные кладки содержат 5—6 яиц. В южном Таджикистане размеры кладки 8—16 яиц (Тугаринов, Козлова, 1945; наши данные). Окраска яиц бледно-палевая, почти полностью выцветающая к концу насиживания,с небольшим количеством бурых крапинок и маленьких пятен.
Масса свежего яйца 25.5—29.5.
Самка насиживает очень плотно, но обычно не позволяет приближаться к ней вплотную. Она способна давать действенный отпор некрупным хищникам, и мне приходилось видеть отснятые японскими орнитологами кадры, на которых показывалось, как насиживавшая самка напала на оказавшегося рядом полоза и убила его, хватая лапами, как хищная птица, и нанося удары лапами и клювом. Есть непроверенные данные о том, что при появлении беспокоящих факторов самка может перетаскивать яйца в другое место. Насиживание продолжается 21—23 дня. Насиживающая самка сильно худеет, теряя до 40 % своего зимнего веса к моменту вылупления птенцов. Она теряет очень много перьев, особенно на брюхе, груди, на внутренней поверхности бедра и голени. В процессе насиживания она отлучается на кормежку дважды в сутки — по утренним и вечерним зорям, оставаясь в гнезде в самое жаркое время дня. Видимо этой необходимостью — находиться на жаре каждый день — и объясняется выпадение у самки столь значительной части перьев.
Вылупление птенцов происходит в гнезде в течение суток. Масса новорожденного птенца 21.5—23. Пеньки маховых перьев появляются у птенцов только на 3-й день. Выводок несколько дней держится в районе гнезда, а затем начинает перемещаться в места с наиболее благоприятными в данное время кормовыми условиями. На 13-й день птенцы могут взлетать на 30 см, в 20-дневном возрасте они способны пролетать 15—30 м над землей, а в 30-дневном — уже осваивают технику вертикального «взрывного» взлета (Тугаринов, Козлова, 1945). Развитие оперения у птенцов, согласно упомянутым авторам, на юге Таджикистана происходит следующим образом. Маховые перья на 4-й день достигают 3 см длины, на 6-й хорошо видны большие кроющие перья крыла и второстепенные маховые, на 14-й день появляются плечевые перья, раскрываются пеньки рулевых. На 17-й день появляются пеньки ювенальных перьев на груди, на 19-й — на надхвостье, в 3-недельном возрасте — на спинной птерилии. В возрасте 25 дней обозначается маленьким бугорком на плюсне шпора, а в 30-дневном возрасте оперяется голова: сначала кроющие перья уха, затем темя, затылок и лоб. В возрасте 40 дней начинается смена ювенальных маховых перьев, хотя сам ювенальный наряд полностью развивается только к 45-му дню жизни. В возрасте 50 дней появляются перья переходного наряда, а на 52-й день — и дефинитивные контурные перья, сначала только на груди. Линька в первый взрослый наряд заканчивается в возрасте 5 мес, в октябре.
Размеры и масса тела у него примерно такие же, как у домашней курицы, но благодаря длинным хвосту и ногам самец выглядит несколько крупнее.
Окраска самца очень яркая, состоит из сочетания красновато-рыжего, черного, белого, золотистого, синего и зеленого цветов, тогда как самка окрашена в скромные сероватые тона с большим количеством коричневых пестрин. В природе фазана невозможно спутать ни с какой другой птицей прежде всего благодаря его длинному хвосту, особенно бросающемуся в глаза у летящей птицы. Фазан очень быстро бегает и легко пробивается в самых густых зарослях, но при близкой опасности взлетает характерным «взрывным» полетом почти вертикально и сразу же развивает большую скорость, хотя пролетает обычно небольшое расстояние. Сам полет типичного-для куриных птиц типа, при котором серии быстрых взмахов чередуются со скольжением на расставленных и загнутых вниз крыльях.
Вопреки существующему мнению, фазан сравнительно часто пользуется полетом, особенно там, где места кормежек, ночевок и водопоя находятся на некотором удалении друг от друга. Те популяции, которые совершают значительные сезонные перекочевки, делают это как с помощью полета, так и пешком, но пока еще никто не наблюдал беспосадочного полета фазана дальше чем на 1 км.
Будучи наземной птицей, фазан тем не менее охотно кормится на деревьях осенью и зимой, а птицы из кавказских популяций ночуют на них и летом. Голос самца — грубый двусложный крик, причем каждый из слогов в свою очередь тоже двусложный, но произносится почти слитно, особенно первый. Чаще всего он слышен в весеннее время, но может издаваться и испуганной птицей при взлете. Чем-то встревоженный, самец издает односложный позыв «кох», повторяемый с небольшими интервалами все время, пока птица беспокоится. Самка очень молчалива и, только будучи вспугнутой, издает при взлете тонкий писк.
Здесь фазаны проводили обычно середину дня, греясь на солнце в защищенных от ветра местах, Кормились они также иа южных склонах — по окраинам сельскохозяйственных полей, поселков, у дорог, заходя даже в пределы крупных городов по речным поймам. На ночь они без труда находили себе убежище в густых зарослях низкорослого бамбука на крутых, опять же южных склонах. Наиболее обычным местом ночевки был нагнувшийся под тяжестью снега бамбук: сиег на нем выполнял роль крыши, а толстая подстилка из сухой травы предохраняла птицу от холода смерзшейся земли. В южном Таджикистане обычными местами ночевок, чаще групповых, также являются густые заросли невысоких тростников, обычно невдалеке от водопоев.
Сезонные перемещения. В большинстве случаев фазан отличается большой привязанностью к одной и той же территории в течение всей своей жизни. Максимальная оседлость особенно характерна для небольших изолированных популяций Закавказья и Средней Азии. На севере ареала (Балхаш, Приамурье) птицы совершают регулярные сезонные перемещения на большие расстояния (иногда до 200 км по прямой) из мест, где устанавливается значительной толщины снежный покров, в малоснежные или совсем бесснежные районы. Это свойственно также и горным популяциям. Сезонные перекочевки совершаются как небольшими группами, так и стаями и даже одиночными птицами. В прибалхашских зарослях такие перекочевывающие стаи сразу же выделялись среди местных фазанов постоянством своего состава, тесной связью птиц в стаях и их совместными перемещениями (Штегман, 1945). Но в целом сезонные перемещения фазанов изучены еще очень плохо, особенно на Дальнем Востоке, где они подчас приобретают большие масштабы.
Смена оперения у взрослых птиц проходит ежегодно в течение лета. Она начинается, по крайней мере у самцов, с замены 1-го первостепенного махового пера. На стадии смены 4-го первостепенного махового начинается активная линька контурного оперения по всему телу птицы, что происходит в начале июня. В конце июня начинается смена второстепенных маховых перьев, сначала от самых внутренних кнаружи, затем от 2-го внутрь таким образом, что последними сменяются расположенные в центре 7-е и 8-е второстепенные маховые, по направлению к которым и идет смена перьев с двух сторон. Рулевые перья сменяются от наружных к центральным парам. Линька перьев крыла и хвоста нередко проходит ассиметрично.
У самок линька задерживается насиживанием, но затем активно протекает примерно по той же схеме, что и у самца. Окончание линьки — в начале сентября.
Подробнее о сокращениях можно прочесть здесь.
Песня
…
Позывка
…
Электроманок
…
Литература
• «Птицы Советского Союза». Дементьев Г.П., Гладков Н.А., Исаков Ю.А., Карташев Н.Н., Кириков С.В., Михеев А.В., Птушенко Е.С. Под общей редакцией Г.П. Дементьева и Н.А. Гладкова. Изд. «Советская наука» Москва. 1952 г. том 4. стр. .
Литература
• Коблик Е.А. 2025. Фауна птиц стран Северной Евразии в границах бывшего СССР: Списки видов.
• IOC World Bird List v15.1 (Updated February 20, 2025) by Frank Gill, David Donsker & Pamela Rasmussen.
• Н.Н. Балацкий. Птицы Северной Евразии. Таксономический список: август 2023 г.
• «Птицы СССР. Курообразные. Журавлеобразные». т.2. Р.Л. Беме, Н.П. Грачев, Ю.А. Исаков, А.И. Кошелев, E.Н. Курочкин, Р.Л. Потапов, А.К. Рустамов, В.Е. Флинт. Ленинград. Издательство «Наука». 1987 г. стр. 119-135.
Сайты
• zmmu.msu.ru
• www.balatsky.ru
• www.worldbirdnames.org
• Avibase by Denis Lepage
• Zoonomen by Alan Peterson
• Taxonomy by John Boyd
• сайт Василия Вишневского
• www.xeno-canto.org
• www.featherbase.info, www.pbase.com, www.macaulaylibrary.org