

Камышевка-барсучок Acrocephalus schoenobaenus – один из наиболее обычных и широко распространённых видов птиц Палеарктики. Его ареал простирается от среднеазиатских пустынь до таёжных рек и тундр Заполярья. В то время как на широте средней полосы и в северных частях ареала камышевки-барсучка её биология изучена достаточно полно (Alker, Redfern 1996; Halupka 1996; Hasselquist, Langefors 1998; Borowiec 1999; Попельнюх 2002; Krol et al. 2002; и др., см. Cramp 1992), сведений об обитании камышевки-барсучка в степной зоне довольно мало. Между тем, они представляют несомненный интерес для выявления степени экологической пластичности вида, а также возможных причин его эволюционного успеха. Настоящая работа призвана восполнить этот пробел. Исследование гнездовой биологии камышевки-барсучка мы проводили в целях получения необходимой основы для изучения социального поведения этого вида и его роли в многовидовом сообществе околоводных воробьиных птиц.
Методы, район наблюдений
Наблюдения проводили в Восточном Приазовье 25 апреля – 22 июня 2001,, 11 апреля – 28 июня 2002 и 21 апреля – 25 июня 2003 вблизи хутора Сладкий Лиман Каневского района Краснодарского края (46°с.ш., 39° в.д.). Птиц наблюдали на предварительно размеченных и закартированных контрольных участках. Камышевок отлавливали паутинными сетями, метили стандартными алюминиевыми и цветными пластиковыми кольцами, отдельные участки оперения красили театральным гримом. На контрольных участках осуществляли целенаправленный поиск гнёзд. Для всех построек (n = 64) подробно описаны биотоп, в котором они располагались, характер крепления, гнездовой материал, для большинства сделаны стандартные промеры рулеткой и штангенциркулем, прослежены сроки строительства, откладки яиц, вылупления и вылета птенцов, успешность размножения. Данные о питании птенцов основаны преимущественно на наблюдениях за собирающими корм взрослыми птицами.
Исследования проводили на берегах реки Челбас и на акватории образуемого ею лимана Малого Кущеватого. Большая часть акватории Челбаса занята плавневой растительностью – тростником Phragmites australis и рогозом узколистным Typha angustifolia. Такие же заросли развиты по краям лимана. К водоёмам прилегают поля пшеницы Tricium aestivum, сахарной свёклы Beta vulgaris, люцерны Medicago sativa, подсолнечника Helianthus annuus, кукурузы Zea mays и других культур. Между полями и по берегам плавней высажены лесополосы из белой акации Robinia pseudoacacia с травяным ярусом из пырея Elytrigia pseudocaesia.
По берегам плавней, по сырым участкам среди полей и по берегам мелиоративных канав имеются участки, занятые тростником, клубнекамышом Bolboschoenus maritimus, различными злаками (Calamagrostis epigeios, Elytrigia pseudocaesia, Poa sp., Beckmannia eruciformis и др.), бурьяном и разнотравьем (Conium maculatum, Artemisia vulgaris, Artemisia austriaca, Cirsium ucrainicum, Sonchus arvensis, Lactuca tatarica, Epilobium hirsutum и др.).
Результаты
Прилёт
Появление первых птиц отмечено 14 апреля 2002. На контрольном участке в 2002 году первые территории заняты 16 апреля. Массовый прилёт камышевок-барсучков наблюдали 23-26 апреля 2002 и 24 апреля 2003. Последние пролётные птицы в плавнях отмечены 12 мая 2003. Последние самцы на контрольном участке в 2002 году появились 6 июня. Самки прилетают на места размножения позже самцов. В 2002 году первая самка отмечена 25 апреля; в предыдущий год самки прилетели несколько раньше: 25 апреля 2001 уже наблюдали строительство гнезда.
Биотопическое распределение
Пролётные птицы держатся в тростниках, – как по берегам водоёмов, так и в глубине плавней. Пролётные самцы поют и могут останавливаться на несколько дней, охраняя территории, в местах, не пригодных для размножения. Так, один самец держался в густых тростниках на берегу плавней 14-24 апреля 2002. В зарослях рогоза камышевки-барсучки не отмечены.
Самцы камышевки-барсучка, только прилетевшие на места гнездования, также держатся в зарослях тростника. Однако, освоившись с местностью, они занимают территории по краям зарослей, часто в тех местах, где тростник растёт вперемешку с вейником, пыреем и другими злаками. Участки с такой растительностью в месте проведения наших работ занимали до 80% самцов (описана 81 территория). Отдельные самцы селятся по краям залитых водой клубнекамышовых полян, окружённых тростником, в тростнике со злаками (без густых куртин вейника) или в местах без тростника, но с бурьяном из сухих стеблей болиголова или полыни, также со злаками. Некоторые территории располагались в зарослях зелёного болиголова и разнотравья. Один самец рекламировал территорию на поле пшеницы, в месте, где над ней возвышались зелёные стебли бодяка. На значительном удалении от воды, в зарослях, не граничащих с водоёмами (постоянными или временными), камышевки-барсучки не живут. Хотя территории многих самцов (около 40 %) и не включают увлажнённые участки, птицы регулярно летают к воде для сбора корма или для отдыха (в жаркие часы). Одной из важных характеристик территории является наличие на ней элементов растительности, выдающихся над основной поверхностью зарослей: в условиях Приазовья – высоких сухих стеблей тростника, реже – сухих стеблей болиголова, полыни или горца Polygonum sp., стеблей бодяка, поросли ясеня Fraxinus ornus. Такие стебли самцы используют как песенные присады в период рекламирования участков.
Места, выбираемые самками для размещения гнёзд, в общих чертах сходны с теми, которые рекламируют самцы. Описание расположения 64 гнёзд показало, что 71.9 % от общего числа наблюдавшихся самок отдали предпочтение зарослям с преобладанием тростника и вейника, 14.1 % – зарослям тростника и клубнекамыша, 4.7 % – зарослям тростника и пырея, 3.1 % – зарослям тростника, вейника и болиголова, 3.1 % – зарослям тростника с мятликом, бекманнией и другими травами, 1.6 % – зарослям вейника, мятлика, чины и других трав и 1.6 % – зарослям пшеницы и бодяка. Только 7 гнёзд (10.9 %) располагались над влажной почвой, остальные были размещены в сухих местах; 42 гнезда (66.0 %) были удалены от воды более чем на 15 м.
В Приазовье камышевка-барсучок селится в тесном соседстве с индийской камышевкой Acrocephalus agricola, камышовой овсянкой Emberiza schoeniclus, соловьиным сверчком Locustella luscinioides, другими воробьиными птицами. Сведения о составе и динамике сообществ околоводных воробьиных птиц Приазовья частично опубликованы (Иваницкий и др. 2002; Marova-Heinbub et al. 2003; Квартальнов 2005).
Биология размножения
Самцы камышевки-барсучка формируют довольно плотные поселения. В 2001 году на контрольном участке площадью 2 га гнездились 19 самцов; в 2002 году на 4 га – 22 самца. Кроме гнездившихся, в поселениях присутствовали самцы, которым не удалось привлечь самок (в 2002 году – 4 самца). По причине формирования полигамных трио, число гнёзд (не считая повторных) превышает число размножающихся самцов.
Самка начинает строительство гнезда спустя сутки после образования пары, реже – через 2-3 дня. Самец лишь сопровождает её. Строительство начинается с сооружения небольшой платформы, которая представляет собой фрагменты листьев злаков, корешки тростника, пух тростника, паутину и разную растительную ветошь, уложенные между вертикальных стеблей тростника, на листьях злаков или клубнекамыша на некоторой высоте над землёй. Самки барсучка никогда в начале строительства не делают оплёток вокруг вертикальных стеблей, поэтому заготовка гнезда должна иметь опору снизу или располагаться в месте, где вероятность расхождения вертикальных стеблей, поддерживающих заготовку, невелика. После сооружения основания самка строит наружные стенки. Основным материалом стенок служат листья тростника, вейника, других злаков и клубнекамыша. В процессе строительства в стенки гнезда включаются стебли и листья растений, на которых устроено гнездо, что придаёт постройке дополнительную прочность. Затем самка укладывает внутренний слой, состоящий из рыхлого мягкого материала – пуха тростника и других растений, кусочков метёлок тростника с пухом, шерсти, перьев птиц. Этот слой уплотняется в процессе роста птенцов, расширяя полость лотка. Сам лоток самка выстилает сухими веточками метёлок тростника и сухими стебельками других растений. Продолжительность строительства гнезда (от начала до откладки 1-го яйца) составляет 5-7, реже до 9 дней.
Осмотренные нами 59 законченных построек включали следующие материалы: корешки тростника (59.3 % от числа всех гнёзд), сухие листья тростника (32.2 %) и вейника (96.7 %), сухие веточки метёлок тростника (100 %), кусочки метёлок тростника с остатками пуха (не менее 18.7 %), пух тростника (71.2 %), сухие стебельки подмаренника (44.1 %), кусочки метёлок и сухие стебельки вейника (27.1 %), сухие листья клубнекамыша (22.0 %), сухие стебельки осота (15.3 %), сухие листья осота (10.2 %), перья птиц – кряквы Anas platyrhynchos, фазана Phasianus colchicus, соловьиного сверчка Locustella luscinioides – 10.2 %, шерсть енотовидной собаки Nyctereutes procyonoides – 8.5 %, паутина и коконы пауков и насекомых – 15.3 %. Реже встречались сухие стебельки горца, полыни, кривоцвета Lycopsis arvensis и мятлика, колоски, стебельки и сухие листья бекманнии, влагалища и стебельки тростника, сухие листья пшеницы и бодяка, пух бодяка, осота и чертополоха. Компоненты, встреченные лишь в некоторых гнёздах, иногда составляли основу этих построек. Так, в единственном гнезде, где были отмечены сухие листья пшеницы, они служили главным материалом для построения наружных стенок.
Растением, дававшим преимущественную опору для гнезда, в 45.3 % случаев оказывался сухой и зелёный вейник, в 32.8 % – сухой тростник, в 10.9 % – сухой клубнекамыш, в 3.1 % – пырей, в 1.6 % – полынь, в 1.6 % – ситник, в 1.6 % – горец, в 1.6 % – пшеница (по описанию 64 гнёзд). Часто опорой одновременно служили стебли и листья нескольких видов трав – в основном тростника и других злаков. Кроме перечисленных, среди растений, поддерживающих гнёзда, отмечены осот (сухие стебли), бодяк (зелёные растения), зелёные стебли подмаренника, зелёные стебли тростника.
Готовые гнёзда представляли собой сооружения чашевидной формы, с толстыми стенками. Параметры гнёзд (n = 59, в скобках даны минимальное и максимальное значения показателей), мм: внешний диаметр 88.2±7.3 (74-105); диаметр лотка 56.2±5.4 (46-74); высота гнезда 77.6±12.2 (58.5-107); глубина лотка 45.0±6.2 (30-56). Высота гнёзд над поверхностью земли составляла 18.8±10.3 (0-54) см.
Из 15 случаев, когда гнёзда были осмотрены в день перед откладкой первого яйца, в семи (46.7 %) самка ещё продолжала строительство. В четырёх случаях (26.7 %) гнездо оставалось достроенным, но пустым 1 день, в трёх случаях – 2 дня, в одном случае – 3 дня.
Самки откладывают яйца, как правило, каждый день, в утренние часы. Процесс откладки яиц подробно прослежен в 16 гнёздах. В трёх случаях перерыв между откладкой двух последующих яиц составлял 2 сут: в одном гнезде – между откладкой 3-го и 4-го яиц (полная кладка 4 яйца), во втором – между откладкой 4-го и 5-го (полная кладка 5 яиц), в третьем – между откладкой 5-го и 6-го (полная кладка 6 яиц).
Полная кладка состоит в среднем из 5.37±0.64 яиц (n = 38). Найдены 2 кладки из 4 яиц (5.3 %), 21 – из 5 яиц (55.3 %), 14 – из 6 яиц (36.8 %), 1 кладка – из 7 яиц (2.6 %). Один раз отмечена инкубация ещё не законченной кладки с 3 яйцами (полная кладка в этом гнезде состояла из 6 яиц), в большинстве других гнёзд плотное насиживание начиналось со дня откладки предпоследнего или последнего яйца.
Участие самца в насиживании кладки нами не отмечено, не наблюдали и кормление самки самцом. В то же время, самец регулярно посещает гнездо, помогая отгонять от него хищников. Продолжительность инкубации с момента окончания откладки яиц до вылупления первого птенца составляет 11-12 дней, до вылупления последнего – 12-13 дней. Последний птенец появляется на свет либо в тот же день, что и первый, либо на следующие сутки.
Самка обогревает птенцов первые дни после вылупления и защищает их от солнца. Носят корм в гнездо оба партнёра, или (во вторых гнёздах полигамных самцов) только самка. В питании птенцов отмечены пауки, нимфы кузнечиков (Tettigoniidae) и сверчков (Gryllidae), стрекозы сем. Coenagrionidae (имаго), подёнки сем. Baetidae (имаго), тли сем. Aphidae, бабочки (имаго сем. Tineidae, Noctuidae и др.; гусеницы), различные двукрылые (имаго, Culicidae, Tabanidae, Syrphidae и дрю, в том числе – небольшие мошки, ближе не определённые). В поисках корма птицы могут улетать на несколько десятков метров от гнезда.
Вылет птенцов подробно прослежен для 7 гнёзд. Старшие птенцы, если гнездо не подвергается опасности, находятся в нём от 11 до 15 дней. Как правило, все птенцы покидают гнездо одновременно, так что младшие оставляют его в более раннем возрасте – 10-14 сут. Из одного гнезда младший птенец вылетел на день позже, чем остальные. Даже у птенцов, которые покидают гнездо по достижении 15-сут возраста, маховые перья ещё не достигают длины, необходимой для нормального полёта, так что первое время слётки держатся в окрестностях гнезда. Многие птенцы не удаляются от гнезда на значительное расстояние и позже, вплоть до обретения ими самостоятельности, поскольку самец, принимающий участие в кормлении, не улетает далеко со своей территории. Самка либо перестаёт кормить слётков, приступая ко второму циклу гнездования, либо, продолжая кормление, остаётся на территории вместе с самцом. Выводок, выкармливаемый одной самкой, вскоре после вылета из гнезда покидает его окрестности.
Наибольший отмеченный нами период кормления слётков составлял 19 дней после их вылета и, соответственно, 33-34 дня после вылупления (в этом случае их кормил один самец, а его самка насиживала вторую кладку). Слётки, покинувшие другое гнездо, на 20-й день после вылета кормились самостоятельно.
Успешность гнездования зависит от численности хищников. В 2001 году только в 3 из 35 найденных гнёзд вылетели птенцы, ещё 3 гнезда к моменту окончания наблюдений содержали законченные кладки. Всего 23 гнезда (67.6 %) погибли из-за разорения хищниками на стадиях откладки яиц и инкубации. В одном из этих гнёзд самка бросила кладку после того, как было повреждено одно из яиц. Ещё два гнезда (5.9 %) разорены после вылупления птенцов. Заготовки 3 гнёзд брошены недостроенными, а строившие их самки покинули территории партнёров. Частота появления «болтунов» (неоплодотворённых яиц) в гнёздах, где вылупились птенцы, составила 6.7 % (2 из 30 яиц). Из 110 яиц, судьба которых была прослежена, вылупились 28 птенцов (25.5 %), 74 (67.3 %) съедены хищниками, 6 (5.5 %) брошены самкой, 2 (1.8 %) оказались «болтунами». Из 28 птенцов вылетели 15 (53.6 %), 13 (46.4 %) съедены хищниками.
В 2002 году птенцы благополучно покинули 18 из 29 гнёзд. В двух гнёздах птенцы к моменту окончания наблюдений ещё не были готовы к вылету. Разорены хищниками на стадиях откладки яиц и инкубации 4 гнезда (13.8 %), 3 гнезда (10.3 %) разорены после вылупления птенцов, 2 кладки (6.9 %) брошены самками, одна из них – после гибели зародышей (самка насиживала кладку не менее 12 дней; вскрытие яиц показало, что зародыши в них погибли на ранних стадиях развития).
Из 109 яиц в кладках, прослеженных до вылупления птенцов, 3 (2.9 %) оказались «болтунами», ещё одно было повреждено дроздовидной камышевкой Acrocephalus arundinaceus. Всего из 140 найденных в 2002 году яиц барсучка вылупились 112 птенцов (80.0 %), 8 (5.7 %) погибли по вине хищников, 10 (7.1 %) брошены самками (в том числе после гибели зародышей), 3 (2.1 %) оказались «болтунами», одно (0.7 %) погибло из-за дефекта в скорлупе (лакуна, лишённая известковой оболочки и затянутая лишь плёнкой). Из 117 птенцов благополучно вылетели 96 (82.1 %), 13 (11.1 %) были съедены хищниками, 8 (6.8 %) оставались в гнёздах к моменту окончания наблюдений.
На основании характера повреждений скорлупы установлено участие в разорении гнёзд мыши-малютки Micromys minutus и дроздовидной камышевки. Часть кладок, по-видимому, разорили сороки Pica pica. В гибели большинства гнёзд с птенцами, скорее всего, повинны ласки Mustela nivalis. Возможно, гнёзда разоряли также обыкновенные ужи Natrix natrix и волчки Ixobrychus minutus. В 2002 году, когда успешность размножения оказалась высокой, на площадке не отмечены мыши-малютки и волчки, ужи были редки, ласки и сороки стали появляться с конца мая и в небольшом числе. Такая ситуация была обусловлена положением поселения, помещавшегося на удалении от лесополос и зарослей прибрежной растительности.
Данными о выживаемости слётков мы не располагаем. Наибольшую опасность для них может представлять хищничество ласки: 7 мая 2001 наблюдали удачную охоту этого зверя на взрослую камышевку-барсучка. Ласка схватила птицу, когда та кормилась в тростниках у земли, на нейтральном участке, который использовали для сбора корма разные особи поселения. Из пернатых хищников, представляющих опасность для камышевок-барсучков, в районе исследований обычны чеглок Falco subbuteo, обыкновенная пустельга Falco tinnunculus и болотный лунь Circus aeruginosus.
Репродуктивный потенциал камышевок-барсучков в Приазовье существенно увеличивается за счёт склонности многих самцов к полигамии, наличие которой показано наблюдениями за индивидуально мечеными птицами. Из 22 самцов, гнездившихся на контрольной площадке в 2002 году, 7 (31.8 %) после окончания откладки первой самкой яиц и начала инкубации рекламировали вторые территории, на периферии тех, которые опевали ранее (Квартальнов 2005). Привлекли вторых самок 4 самца (18.2 %). В 2001 году привлекли вторую самку 5 из 19 гнездившихся на контрольном участке самцов (26.3 %). Вторые самки приступали к откладке яиц спустя 10-15 дней после первых самок тех же самцов (n = 4). Полные кладки в гнёздах вторых самок полигамных самцов насчитывали 5 яиц (n = 3).
В тех случаях, когда первые самки гнездятся успешно, самцы после начала инкубации в гнёздах вторых самок оставляют последних, не принимая участия в защите гнезда, кормлении птенцов и слётков. Если после гибели гнезда первая самка покидает территорию, самцы поддерживают тесные отношения (защита от хищников, кормление птенцов и слётков) со вторыми самками. Одна вторая самка после гибели гнезда первой самки и успешного вылета птенцов в своём гнезде сделала попытку загнездиться второй раз с тем же партнёром.
Часть особей приступают ко второму циклу размножения. В 2001-2002 годах построили вторые гнёзда 5 из 20 пар, чьи птенцы успешно вылетели (25.0 %) (в расчёт не включены вторые самки полигамных самцов, выкармливавшие птенцов в одиночку). Во всех случаях, которые удалось точно проследить (n = 3), состав пар сохранялся. К постройке вторых гнёзд самки приступали в дни вылета слётков. Откладка яиц во вторых гнёздах начиналась спустя 33-37 дней после начала откладки яиц в первых гнёздах, и спустя 3-5 дней после вылета птенцов (n = 3). Полные кладки в гнёздах второго цикла размножения насчитывали 5 яиц (n = 3).
После разорения гнёзд самки в большинстве случаев покидают территорию самца. Так поступили все самки камышевки-барсучка, потерявшие гнёзда на контрольном участке в 2002 году. В 2001 году 4 из 21 самок, потерявших гнёзда (19.0 %), сделали попытку загнездиться на прежней территории во второй раз. Два повторных гнезда были брошены недостроенными, так как партнёры этих самок к моменту потери гнёзд ухаживали за своими «вторыми» самками. Остальные повторные гнёзда были разорены.
После потери единственного гнезда самцы возобновляли рекламирование территорий. Привлечь вторую самку после потери первого гнезда удалось 3 самцам из 13, оказавшихся в такой ситуации на контрольном участке в 2001 году (23.1 %). В 2002 году ни один из самцов после гибели гнезда не смог образовать пары.
Сроки гнездования
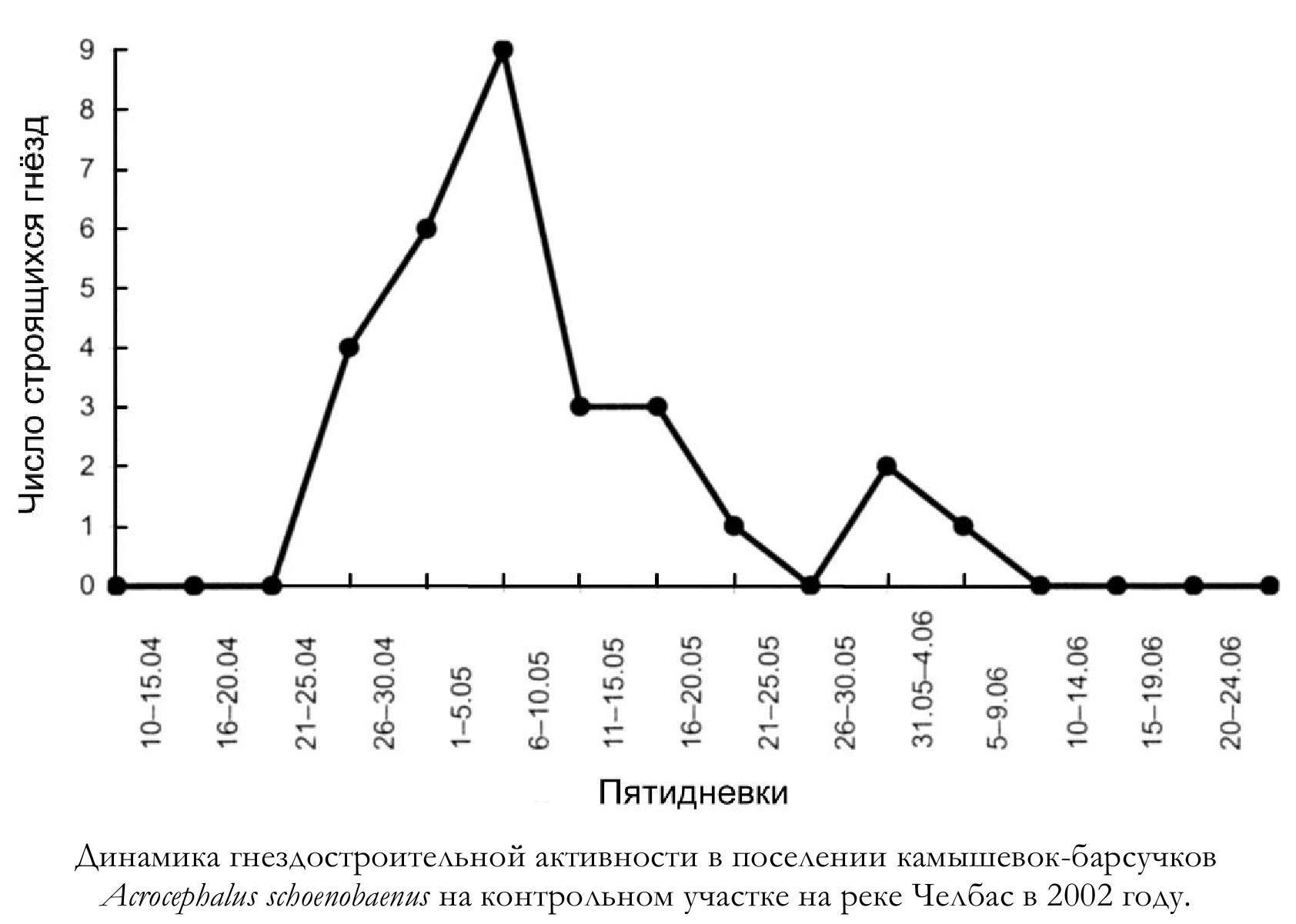
Динамика строительства гнёзд в поселении, изученном в 2002 году, показана на рисунке. Начало откладки яиц в первых гнёздах относится к 5 мая – 31 мая, средняя дата – 12 мая (n = 20). Откладка яиц во вторых гнёздах полигамных самцов начата 18-21 мая, в среднем 20 мая (n = 4). Кладки второго цикла гнездования начаты 8-10 июня, в среднем 9 июня (n = 3).
В 2001 году откладка яиц в первых обнаруженных гнёздах начата 4 мая. Размножение в изученном поселении растянуто из-за того, что многие гнёзда были разорены, и самки строили повторные. Некоторые самцы, довольно поздно приступив к гнездованию, по-видимому, образовали пары с самками, ранее покинувшими других партнёров после разорения гнёзд. Откладка яиц в самом позднем из обнаруженных нами гнёзд начата 17 июня 2001.
Заключение
В основных чертах биология камышевки-барсучка в Приазовье близка к тому, что известно для других точек его ареала. Величина кладок в Приазовье практически не отличается от показателей Германии, Польши, Финляндии и северо-запада России (Cramp 1992; Halupka 1996; Попельнюх 2002; Фетисов и др. 2002), однако значимо больше (Р = 0.028), чем в популяции Британских островов (Bibby 1978).
Как и в других локальных популяциях (Попельнюх 1996а; и др.), успешность размножения варьирует по годам, отход яиц и птенцов в Приазовье в годы с относительно низким обилием хищников сходен с тем, который наблюдается в Англии (Cramp 1992), Польше (Halupka 1996) и Приладожье (Попельнюх 1996а, 2002).
Ко второму циклу размножения приступает четверть самок, успешно выкормивших птенцов. Это значительно выше, чем в Англии, Приладожье и Польше, где в большинстве изученных популяций ко второму циклу гнездования приступают единичные пары (Borowiec, Lontkowski 1988; Cramp 1992; Попельнюх 19966, 2002; Krol et al. 2002). Однако для одной локальной популяции на северо-западе Англии указана высокая степень полициклии – более 90 % по наблюдениям за 18 парами (Alker, Redfern 1996). В тесной связи с высокой частотой второго цикла гнездования находится, по-видимому, высокая частота полигамии. Число самцов, пытающихся привлечь вторых самок, в Польше такое же, как и в Приазовье – 32.5 %, в среднем по трём годам (Borowiec, Lontkowski 1988), однако успешно привлекают вторых самок только 7.3 % самцов, по другим данным – до 10 % (Borowiec 1999). Несмотря на сходное число самцов, пытающихся привлечь вторых самок, немногим самцам удаётся это сделать и в Англии – не более чем 7.7 % (Alker, Redfern 1996; Buchanan, Catchpole 2000). Невысока частота полигамии в ряде других участков ареала (Cramp 1992; Leisler, Catchpole 1992; Попельнюх 2002). Таким образом, у камышевки-барсучка число случаев полигамии зависит не от поведения самцов, но от соотношения полов среди способных к размножению птиц. Допустимо предположить, что «избыток» самок в популяции Приазовья формируется за счёт птиц предыдущего года рождения, принадлежащих к выводкам второго цикла гнездования, поскольку известно, что у многих птиц самки достигают половой зрелости раньше, чем самцы (Паевский 1985).
Самцы из поздних выводков, вероятно, не гнездятся во второе лето жизни: не исключено, что к ним принадлежат птицы, занимающие территории в начале июня и в размножении не участвующие (наши данные по Приазовью). Следует отметить, что относительно высокая частота полигамии указана для шведской популяции барсучка (Hasselquist, Langefors 1998). Там 38 % самцов пытаются привлечь вторых самок, 19 % становятся полигамными. Вторые кладки в Швеции наблюдаются только в отдельные годы, но соотношение полов среди размножающихся птиц сдвинуто в сторону преобладания самок, поскольку 27 % всех территориальных самцов остаются холостыми.
Таким образом, популяция камышевки-барсучка в Приазовье находится в благополучном состоянии. Несмотря на опустошительное воздействие хищников на отдельные поселения, уровень воспроизводства камышевки-барсучка достаточен для поддержания стабильной численности. В то же время, камышевка-барсучок в Приазовье является относительно немногочисленным видом, что определяется незначительной площадью подходящих для неё биотопов. Избыток молодых птиц, вероятно, устраняется за счёт перераспределения их по ареалу.
Литература
• Иваницкий В.В., Марова И.М., Квартальное П.В., Маркитан Л.B. 2002. Сравнительный анализ населения камышевок (Acrocephalus, Sylviidae, Aves) на лиманах Краснодарского края и на степных озёрах Калмыкии // Тр. Тебердинского заповедника 31: 125-130.
• Квартальнов П.В. 2005. Структура сообщества камышевок юга России. Автореф. дис. … канд. биол. наук. М.: 1-24.
• Паевский В.А. 1985. Демография птиц // Тр. Зоол. ин-та АН СССР 125: 1-286.
• Попельнюх В.В. 1996а. Бициклия размножения камышевок рода Acrocephalus в юго-восточном Приладожье // Материалы 2-й конф. молодых орнитологов Украины. Черновцы: 143-145.
• Попельнюх В.В. 1996б. Фенология размножения, размеры и судьба кладок камышевки-барсучка в юго-восточном Приладожье // Материалы 2-й конф. молодых орнитологов Украины. Черновцы: 146-147.
• Попельнюх В.В. 2002. Экология камышевок рода Acrocephalus в юго-восточном Приладожье // Тр. С.-Петерб. общ-ва естествоиспыт. (Сер. 4) 87: 1-144.
• Фетисов С.А., Ильинский И.В., Головань В.И., Фёдоров В.А. 2002. Птицы Себежского Поозерья и национального парка «Себежский» // Тр. С.-Петерб. общ-ва естествоиспыт. (Сер. 6) 3, 2: 1-128.
• Alker P.J., Redfern C.P.F. 1996. Double brooding and polygyny in Sedge Warblers Acrocephalus schoenobaenus breeding in north-west England // Bird Study 43: 356-363.
• Bibby CJ. 1978. Some breeding statistics of Reed and Sedge Warblers // Bird Study 25: 207-222.
• Borowiec M. 1999. Biologia i ekologia legowa podwrociawskiej populacji rokitniczki (Acrocephalus schoenobaenus). Zachowanie rozrodcze i strategie ewolucyjne samcцw i samic // Acta Univ. Wratislaviensis, Prace Zoologiczne 33: 1-92.
• Borowiec M., Lontkowsky J. 1988. Polygyny in the Sedge Warbler Acrocephalus schoenobaenus // Vogelwelt 109, 5/6: 222-226.
• Buchanan K.L., Catchpole C.K. 2000. Extra-pair paternity in the socially monogamous Sedge Warbler Acrocephalus schoenobaenus as revealed by multilocus DNA fingerprinting // Ibis 142: 12-20.
• Cramp S. (ed.). 1992. The Birds of the Western Palearctic. Oxford Univ. Press, 6: 1-728.
• Halupka L. 1996. Breeding ecology of the Sedge Warbler Acrocephalus schoenobaenus in the Biebrza Marshes (SW Poland) // Ornis hungarica 6: 9-14.
• Hasselquist D., Langefors Д. 1998. Variable social mating system in the Sedge Warbler, Acrocephalus schoenobaenus // Ethology 104: 759-769.
• Krol W., Solarz W., Zajac T. 2002. Breeding biology of the sedge warbler Acrocephalus schoenobaenus in the river Nida wetlands (Poland) // Biologia (Bratislava) 57: 621-629.
• Leisler В., Catchpole C.K. 1992. The evolution of polygamy in European reed warblers of the genus Acrocephalus: a comparative approach // Ethol., Ecol. & Evolution 4: 225-243.
• Marova-Kleinbub I., Markitan L., Kvartalynov P., Ivanitskii V. 2003. The structure of reed warbler (Acrocephalus spp.) community on the estuaries of Southern Russia // Vogelwarte 42: 44.
Второе издание. Первая публикация в 2005 г. Квартальнов П.В. 2005. Гнездовая биология камышевки-барсучка в Восточном Приазовье // Орнитология 32: 71-77.
Источник
Русский орнитологический журнал 2017, экспресс-выпуск 1547. стр. 5557-5567.