

Семейство: Фазановые Phasianidae
Род: Каменные куропатки Alectoris
Вид: Alectoris chukar (Gray, JE 1830)
Протоним: Perdix Chukar.
Синоним: Caccabis saxatilis, Alectoris graeca, Perdix chukar, Alectoris kakelik, Tetrao kakelik.
Немецкое название: Chukarhuhn
Испанское название: Perdiz chúcar.
Казахское название: Кекілік
Украинское название: Кеклик.
Другие названия: каменная куропатка.
Taxonomic Serial Number : 175908 | Global Biodiversity Information Facility : 2474029.
Список ПОДВИДОВ вида Кеклик Alectoris chukar (Gray, JE 1830)
‐ . — Alectoris chukar kleini (Hartert, EJO 1915)
‐ Восточноиранский кеклик — Alectoris chukar koroviakovi (Zarudny, NA 1914)
‐ Кызылкумский кеклик — Alectoris chukar subpallida (Zarudny, NA 1914)
‐ Тяньшаньский кеклик — Alectoris chukar falki (Hartert, EJO 1917)
‐ Джунгарский кеклик — Alectoris chukar dzungarica (Sushkin, PP 1927)
‐ Кашгарский кеклик — Alectoris chukar pallida (Hume, AO 1873)
‐ Кара-Корумский кеклик — Alectoris chukar pallescens (Hume, AO 1873)
ОСНОВНОЕ ОПИСАНИЕ ПО: Кеклик — Alectoris chukar (Gray, JE 1830)
Окраска. Взрослые самец и самка. Оперение кеклика очень красиво: верхняя сторона тела дымчато-серая с винным оттенком на передней части спины и части кроющих крыла. От лба через глаз к уху идет черная полоса, которая окаймляет желтоватое горло. Кроющие уха рыжие. В углах рта и в основании подклювья черные пятна. Первостепенные маховые бурые, на наружных опахалах имеются продольные охристые пятна, образующие на сложенном крыле светлую предвершинную полоску. Брюхо и нижние кроющие хвоста охристо-желтые. Рулевых 14, цвет их каштаново-рыжий. У средних пар в основании пера развит дымчато-серый цвет. Перья на боках туловища с отчетливо выраженными черными и коричневыми .поперечными полосами.
Лапы, клюв и кольцо вокруг глаз красные. Половой диморфизм в окраске оперения отсутствует.
Ювенальный наряд (самцы и самки). Верх тела серовато-бурый, со светлыми треугольными вершинными пятнами на перьях. Нижняя сторона тела более светлая с неясными поперечными бурыми полосами. Верх головы («шапочка») охристый. Клюв черный, радужина бурая, ноги начинают приобретать красный цвет.
Пуховой наряд. Верх головы (лоб, темя, затылок) коричневобурый, щеки и пятно за глазом сливочно-охристые. От глаза к затылку проходит узкая черная полоска, ограничивающая «шапочку». Спина пестрая, темно-коричневый пух образует 3 продольные полоски более интенсивной окраски, чем верх головы. Между ними находятся участки, покрытые длинным серовато-серебристым пухом. На крыле удлинненные слегка охристые пушинки. Подбородок, горло и бока шеи почти белые со слабым желтоватым оттенком. Остальная часть нижней стороны тела покрыта длинным белым пухом с оттенком сливочного цвета. Клюв у вершины желтоватый, у основания темный. Яйцевой зуб белый. Ноги слегка розоватые.
Строение и размеры. Крылья тупые, закругленные, хвост умеренной длины, слегка закруглен. Самцы отличаются от самок наличием шпор. Визуальное определение добытых на юго-востоке Казахстана 329 кекликов с последующим вскрытием и установлением пола показало, что из 191 самца пол по наличию шпоры был верно определен у 187 особей (97.8 %) и только 4 птицы со шпорами оказались самками. В то же время из 138 самок пол по отсутствию шпоры был правильно установлен у 125 птиц (90.6 %), а 13 птиц без шпор оказались самцами. Среди 56 изученных в этом отношении взрослых самок шпоры встречены у 10, причем у 7 они оказались только на одной из лап.
Длина крыла самцов 152—175, самок 142—162. Длина хвоста у самцов и самок 80—90, плюсны 43—47. Масса самцов составляет 450—700, самок 360—550.
Суточная активность, социальное поведение. Суточная активность кеклика четко разграничена на 2 периода — дневной и ночной. В светлый период суток птицы бывают особенно деятельными в утренние и вечерние часы. Летом с восходом солнца кеклики часто встречаются на кормежке, а в жаркие часы дня отдыхают в тени кустов или скал. С наступлением вечерней прохлады активность их вновь возрастает и кормясь, они постепенно поднимаются в верхние части склонов, где и ночуют. Выпадание дождя сильно снижает активность птиц, и они пережидают его в кустарниках, а с прекращением непогоды кормятся вблизи этих мест.
Характерной чертой жизни кекликов в конце лета и в начале осени является регулярное посещение водопоев. С рассветом стайки спускаются к родникам и речкам, чаще преодолевая расстояние по воздуху. Местами водопоев служат открытые участки берегов речек, родников или полого спускающиеся к воде осыпи. В жаркие дни часто устраиваются на отдых в ближайших к водопою кустах, где купаются в пылевых ваннах, напоминающих лотки гнезд без строительного материала.
Кеклики — общественные птицы и большую часть года проводят в стаях. Лишь в период размножения, да й то не все, держатся парами.
По окончании брачного периода особи, не участвующие в насиживании кладок и воспитании молодняка, объединяются в стаи. После вылупления птенцов выводки живут или отдельными стайками, или же объединяются в более крупные стаи, которые обычно не отличаются большим постоянством. Например, в конце августа—начале сентября на особенно кормных местах по склонам гор и днищам ущелий нередко встречаются стаи до 100 особей, но потревоженные, они легко распадаются. Стайки взрослых птиц, обособившиеся в июне и состоящие из самок, потерявших кладки, и самцов, не участвующих в насиживании, распадаются лишь к следующей весне, что подтверждается отловом меченых птиц. В таких стайках обычно не бывает больше 8—12 особей.
Хозяйственное значение, охрана. Охота на кекликов в горных районах Кавказа, Средней Азии и Кахастана издавна пользовалась большой популярностью. В 30-х гг. текущего столетия производились и промысловые заготовки этого ценного вида дичи, поступавшего не только на внутренние, но и на внешний рынки. Только через Ленинградскую экспортную базу в 1927—1928 гг. прошло 166.7 тыс. штук (13.6 % от всей обработанной там дичи), в следующую зиму— 198.1 тысяча (17.9 %), а в несколько последующих зим — более 70 тыс. ежегодно, причем максимальное количество птиц за зиму (1930—1931 гг.) составило 233.2 тыс. (Рудановский, Насимович, 1933, — цит. по: Грачев, 1983). По далеко неполным данным, кеклик в 1962—1963 гг. занимал в Казахстане по добыче второе место после серой куропатки, а в 1965 — первое.
В охотничьи сезоны 1962—1965 гг. ежегодно добывалось от 16 до 53 тыс. штук (Кондратенко, Смирнов, 1973). В «урожайные» годы кеклик становится основным видом горной дичи, добываемой в республиках Средней Азии и Казахстана и имеет существенный вес в общем объеме отстреливаемых охотничье-промысловых птиц. Плановые заготовки в настоящее время не проводятся. Многиебезружейные способы добычи, описанные ранее в литературе (Бутурлин, 1932; Наумов, 1931; Попов, 1956), утратили свое значение или неприменимы из-за большого ущерба, причиняемого популяциям. Основная линия использования кеклика в настоящее время представляет собой спортивную ружейную охоту.
Оптимальным временем для охоты на кеклика следует признать вторую половину ноября—первую половину декабря, когда птицы имеют максимальную массу (взрослые самцы 613 г, взрослые самки 504, молодые самцы и самки соответственно 553 и 475 г).
Своеобразная гнездовая биология кеклика способствует поддержанию высокой численности вида в’благоприятные годы и, что особенно важно, быстрому увеличению поголовья после массовой гибели. Учитывая хозяйственное значение кеклика как одного из главных охотничьих объектов гор юга СССР, следует очень бережно относиться к его поголовью. Это подразумевает ведение постоянных учетов численности поголовья, элементарные биотехнические мероприятия во время суровых многоснежных зим (в основном подкормка) и запрещение охоты на сроки не менее 3 лет после особенно суровых зим.


В пределах СССР кеклик распространен по Главному Кавказскому хребту, в Закавказье (включая Талыш), в западном и центральном Копетдаге, Больших Балханах, западном Узбое, по чинку Устьюрта, на п-ве Мангышлак, по возвышенностям Бадхыза и, возможно, по обрывам верхнего течения рек Теджен и Мургаб. Живет в горных поднятиях пустыни Кызылкум (Аристанбельтау, Кюйгеньтау, Актау, Тохтатау), горах Ходжа-Баба, Байсунтау. Населяет горы Памиро-Алая (кроме Памирского нагорья, — см: Потапов, 1966), систему Тянь-Шаня, Чу-Илийские горы, Джунгарский Алатау, Тарбагатай, Саур, Западный и Южный Алтай, Западный Танну-Ола (рис. 13). Северная и северо-восточная границы ареала до конца не выяснены. Есть нахождения этого вида на западном берегу Аральского моря к югу от Кератамака (сборы Бурачека от 21 IV 1924, кол. ЗИН АН СССР) и в Саянах (сборы А.Я. Тугаринова от 24 IX 1908 у с. Тюрботы, 30 км от р. Кемчик; сборы С.И. Снигиревского с р. Абакан III 1936, кол. ЗИН АН СССР). Имеются также экземпляры, полученные от Абаканской заготовительной конторы в январе—апреле 1936 г., которые могли поступать сюда с западного Танну-Ола. До сих пор нуждается в уточнении вопрос об обитании кеклика в Мугоджарах (Зарудный, 1888), Улутау (Павлов, 1934), в окрестностях Семипалатинска и в Семейтау (Хахлов, Селевин, 1928).
Акклиматизирован в горах Крыма.
Численность. Сведения о численности кеклика в различных районах весьма скудны. В Центральном Копетдаге общее количество птиц было определено в 121 000, т.е. 810 выводков (Лаптев, 1936). В последние 20 лет учеты этого вида проводят в основном в охотничьих хозяйствах.
Изучение многолетней динамики численности кеклика на постоянных маршрутах проводилось с 1972 по 1976 гг. в конце августа—середине октября на юго-востоке Казахстана. В хр. Каратау число учтенных птиц на 1 км маршрута менялось за этот период довольно сильно. Результаты учетов показали, что наибольшая численность кеклика отмечена в хребтах Каратау и Алтын-Эмель, где росту численности препятствовали неблагоприятные условия отдельных зим. Наиболее высокая плотность птиц отмечена в Киргизском Алатау, где численность хотя и претерпевала некоторые изменения по годам, но оставалась высокой.
При исключительном разнообразии мест обитания кеклик все-таки отдает предпочтение ущельям с выходами скал и склонами, где каменистые осыпи чередуются с открытыми травянистыми участками, иногда поросшими кустарником. Важную роль играет наличие водопоев (горные реки, ручьи, ключи), в исключительных случаях птицы могут использовать горько-соленую воду. В ряде мест кеклики встречаются в совершенно несвойственных им условиях — например, на равнинах среди песков на значительном расстоянии от ближайших гор (Сержпинский, 1925; Молчанов, 1932; Шнитников, 1949; Ишадов, 1970).
На Кавказе кеклик населяет разнообразные биотопы от сухих склонов гор и предгорий вверх до 3500 м над ур. м., где живет по россыпям вблизи ледников (Сатунин, 1907). По безлесным каменистым участкам гор он спускается почти до равнины, где встречается на гнездовье среди кустарников вместе с турачом. Однако этот вид всегда оказывает предпочтение склонам предгорий или ущельям горных рек с выходами скал и с каменистыми осыпями, где преобладает редкая ксерофитная растительность. Реже занимает луговины северных склонов с арчевым редколесьем, избегая, как правило, сырых участков с богатой растительностью.
В Копетдаге кеклик обитает в небольших ущельях на высотах 500—600 м, иногда поднимаясь до предельных высот хребта (2000 м над ур. м.), но придерживаясь мест, где имеются водопои. Нижняя граница распространения лежит здесь в полынно-эфемеровой полупустыне, а верхняя связана с участками горных ксерофитов. Кроме того, кеклик встречается и среди кустарниковой растительности, распространенной от подножий гор и до их вершин — в арчевниках, зарослях трагакантовых астрагалов, других кустарников, растущих у осыпей и скал, среди зарослей дикого винограда, ежевики и различных плодовых деревьев и кустарников (Федоров, 1949). Населяет он и обрывистые песчаные берега Узбоя в районе пресных озер, где встречается среди саксаула и песчаной акации (Молчанов, 1932).
Наибольших высот в СССР кеклик достигает в Бадахшане. В долине Шахдары верхняя граница его распространения проходит на высотах около 4000 м над ур. м. Здесь кеклик встречается по всему бассейну реки, но численность его невелика; наибольшая плотность отмечена на высотах 2300—2600 м над ур. м. В этих высокогорных условиях птицы ведут оседлый образ жизни и держатся в период гнездования и зимой на каменистых склонах и моренах среди осыпей (Степанян, 1969).
В Тянь-Шане нижний предел вертикального распространения кеклика лежит на высоте 300 м, верхний же достигает 3600 м над ур. м. Здесь птицы живут оседло, совершая лишь небольшие перемещения. Населяют скалистые склоны, покрытые ксерофитным разнотравьем и ягодными кустарниками (вишня, кизильник, жимолость, эфедра). В крупных горных массивах кеклики встречаются от подножий хребтов до 3600 м, а в Киргизском хребте в небольшом числе встречается у вечных снегов (Спангенберг, Судиловская, 1959), но отсутствует однако же на сыртах, в котловине оз. Сонкёль и других высокогорных долинах Центрального Тянь-Шаня (Янушевич и др., 1959).
В Киргизском Алатау кеклик гнездится в зарослях шиповника, на опушках лиственного и хвойного леса, среди скал и осыпей. В осеннее время стаи чаще встречаются в кустарниковых зарослях по склонам гор илн берегам рек. В Таласском Алатау обитает от культурного пояса до субальпийского (1000—3000 м над ур. м.). Населяет сухие каменистые склоны с разреженной травянистой растительностью и редким кустарником. Обычен в арчевом редколесье. Иногда гнездится у подножий гор на участках, лишенных каменистых обнажений с растительностью скорее лугового, чем степного характера (Ковшарь, 1966).
В Зеравшанском, Туркестанском и Гиссарском хребтах населяет скалистые склоны с зарослями кустарников, реже держится на безлесных скалах и осыпях и еще реже на травянистых склонах. Пределы высотного распространения в этих хребтах 1200—3500 м над ур. м.
В хребтах Джунгарского Алатау кеклик наиболее многочислен на высотах 500—1500 м над ур. м., где населяет пустынный, степной и лесо-лугово-степной пояса. В большом количестве встречается в западных отрогах (горы Чулак и Малай-Сары), граничащих с щебнистой пустыней. Обитает в скалистых ущельях с обширными каменистыми осыпями и участками травянистой и кустарниковой растительности, среди кустарников боялыча, эфедры, таволги, курчавки ланцетовидной и боярышника.
На горном плато Малай-Сары кеклик населяет узкие ущелья, выходящие на плато, которое используется под посев зерновых культур. После окончания уборки урожая птицы кормятся оставшимся зерном. На хр. Алтын-Эмель кеклик поднимается до высоты 2000 м над ур. м., где растительность носит некоторый северный колорит и представлена лиственными породами деревьев, образующих вдоль рек густую урему из берез, ив и черемухи. Вдоль ручьев встречаются участки с постоянно зеленым травяным покровом. Высокая численность птиц и небольшие колебания численности имеют место в нижних поясах крупных горных хребтов в пределах до 2000 м над ур. м. Здесь птицы находят наиболее подходящие условия для существования.
В пище кеклика зарегистрировано 311 видов высших растений из 53 семейств, а также мхи и водоросли. Среди животной пищи основу составляют 132 вида насекомых из 36 семейств 10 отрядов. Моллюски встречаются гораздо реже (10 видов), а также, как исключение, поедаются пауки.
По встречаемости, разнообразию видового состава и объему растительная пища в рационе кеклика значительно преобладает над животной. В целом соотношение растительных и животных кормов у кекликов Казахстана выражается следующими цифрами. Только растительная пища обнаружена, у 1076 особей (77.3 %), смешанная у 309 (22.1 %), исключительно животные корма у 8 птиц (0.6 %). Среди растительных кормов можно выделить представителей 9 основных семейств: злаки 35 видов (58.3 % встречаемости), розоцветные 26 (26 %), лилейные 21 (25.6 %), амариллисовые 2 (23.7 %), бурачниковые 26 (15.2 %), зонтичные 14 (13.1 %), сложноцветные 36 (13.1 %), бобовые 31 (12.7 %), гречишные 8 (12.5 %). Остальные семейства в пище кеклика составляют менее 10 % встречаемости. Основные животные корма представлены отрядами прямокрылых (22 вида), жесткокрылых (56 видов) и полужесткокрылых (33 вида).
Все корма кеклика можно разделить на 4 группы: 1 — семена и плоды (в том числе ягоды) травянистых растений и кустарников; 2 — зеленые части растений (листья, стебли, цветы); 3 — подземные части растений (луковицы, клубни, корни, корневища); 4—беспозвоночные животные (насекомые,; пауки, моллюски).
Из всех групп растительных кормов по частоте встречаемости и разнообразию видового состава важнейшее место занимают семена (178 видов 35 семейств). Основу этой группы корма составляют семена злаков (25 видов), бурачниковых (21 вид), крестоцветных (10 видов), лилейных и маревых (по 9 видов). Остальные семейства представлены меньшим числом видов. В течение всего года семена составляют значительную часть пищи, за исключением весенних месяцев, когда сохранившихся после зимы семян остается мало. Особенно большое количество семян поедается во второй половине лета и осенью, когда линяющим птицам после размножения требуется особо питательный и разнообразный рацион. Длительная сохранность семян в значительной степени обеспечивает питание птиц в зимний период.
Из подземных частей растений в рационе кеклика отмечены луковицы, клубни, прикорневые луковички и корни травянистых растений 25 видов (9 семейств), особенно луки, гусиные луки, тюльпаны, но по частоте встречаемрсти на первом месте стоят прикорневые луковицы злаков (луковичного мятлика и луковичного ячменя) и луковицы амариллисовых (иксиорилион и унгерния). Довольно часто в пище кеклика встречаются клубни гераниевых и зонтичных (скалигерия). Зимой добывание глубоко расположенных луковиц и клубней затруднено из-за промерзания почвы, и птицы используют лишь прикорневые луковицы луковичного мятлика.
Зеленые корма довольно разнообразны и представлены 61 видом из 18 семейств высших растений. Основную массу их составляют злаки, а также сложноцветные (15 видов, из них 5 видов полыней, одуванчики), бобовые (13 видов, из них 4 вида астрагалов, ползучий клевер, люцерна, горошки, два вида солодок и др.). Кроме того, встречаются представители маревых, зонтичных, бурачниковых.
Весной (март—апрель) зелень и цветы эфемеров — основная группа корма в рационе кекликов. Среди этой группы особо следует выделить различные виды лилейных (Alleum, Gagea, Tulipa), шафран, луковичный мятлик и иксиолирион.
Ягоды по сравнению с другими группами растительных кормов в рационе играют небольшую роль. Ограниченный видовой состав, относительно низкая встречаемость и короткий период использования делают ягоды явно второстепенным кормом. Всего же в пище кеклика отмечены ягоды 25 видов из 11 семейств. Чаще других поедаются вишня, боярышник, шиповник, кизильник. Ягоды лоховых, пасленовых, тутовых, жимолостных, барбарисовых, виноградных отмечены в питании кеклика на сравнительно небольших территориях. В период массового созревания ягоды в большинстве районов становятся на короткое время основным кормом, вытесняя все другие виды кормов.
Животные корма по своей питательной ценности занимают в рационе кеклика особое место. Например, поедание весной моллюсков способствует восполнению потерь кальция, необходимого для образования скорлупы яиц. Из насекомых наиболее часто поедаются прямокрылые, преимущественно саранчевые (16 видов). Из жесткокрылых чаще других употребляются слоники (23 вида) и листоеды (14 видов), а также другие жуки, точно не определенные. Полужесткокрылые встречаются реже, но по видовому составу они уступают лишь жесткокрылым и в массе поедаются кекликом только в местах скоплений. Например, в ноябре—декабре 1978 г. в Заилийском Алатау (ущелье Жаманты) клопы отмечены в питании 17 особей из 61. Основная масса клопов была представлена Dolicoris penicillatus, который у отдельных птиц составлял 40—85 % содержимого зоба. Здесь же отмечены другие виды клопов: Euridema ornata, Emlethis spp., Corizus hyoscyami, Corioneris histicornis, Sciocoris deltocephalus.
На юго-востоке Казахстана животные корма в пище кеклика (встречаемость 13.4 %) присутствуют с августа по январь (наиболее часто в августе и декабре). Обычно этот вид кормов встречается в качестве небольшого дополнения к растительному, но у отдельных особей может составлять 50—85 % и, как исключение (1 случай), даже 100 % содержимого зоба. Из насекомых осенью особенно часто поедаются саранчевые (30 %), второе место по частоте встреч занимают клопы (27.7 %), которые присутствуют в рационе с октября по декабрь, но в наибольшем количестве отмечаются в ноябре—декабре. Вероятно, птицы находят насекомых в это время в местах их массовых скоплений на зимовках.
Жуки в рационе кеклика отмечены с октября по ноябрь (встречаемость 24.4 %), причем наиболее часто они попадаются в октябре. Муравьи поедаются в небольшом числе с августа по октябрь (12.2 % встреч), моллюски — в сентябре—ноябре.
Летом пища взрослых особей по сравнению с другими сезонами года наиболее разнообразна, обычно в каждом зобу можно обнаружить по 2—3 группы кормов. Животные корма в этот период составляют сравнительно небольшую часть. Лишь в одном случае у насиживающей самки в зобу обнаружено 17 моллюсков Ponsadenia semenowi (30 % от содержимого зоба массой 19 г).
На юго-востоке Казахстана подземные части растений, преобладающие в рационе в августе—сентябре, уступают место в более поздние сроки семенам и зелени — основной пище зимнего сезона. Процесс смены кормов обусловлен не только обилием последних в данное время. Сокращение подземных частей растений в составе рациона наступает задолго до выпадения снега, когда этот вид корма становится менее доступным. Вообще необходимо отметить, что при обилии пищи кеклики проявляют довольно высокую избирательную способность, предпочитая только определенные корма и определенные виды растений.
В разных частях обширного ареала в пределах СССР средняя доля семян, подземных частей растений и зелени в годовом рационе кеклика примерно одинакова и превосходит более чем в 2 раза долю ягод и животных кормов. Вместе с тем, значительные изменения в составе рациона происходят в зависимости от сезона. Так,доля семян и подземных частей растений в течение года изменяется в 4—5 раз, достигая максимума в июле—августе и минимума в апреле. Наоборот, зеленые корма особенно активно потребляются в марте—апреле и наиболее редко — в июне и июле. Встречаемость животных кормов изменяется в 12 раз, с максимумом в июне—августе. Ягоды, при полном их отсутствии в январе, в августе встречены более чем у 40 % особей.
Питание птенцов на территории нашей страны изучено недостаточно. Только в Джунгарском Алатау было определено содержимое зобов и желудков 36 птенцов в возрасте от 2 дней до 1 месяца (Остапенко, 1958).
Корма птенцов в первый месяц жизни оказались здесь весьма разнообразными. В основном поедались насекомые (саранчовые, щнтники, певчие цикады, цикадки, коровки, златки, долгоносики, чернотелки, пластинчатоусые, мухи, муравьи), а также моллюски. Среди растительных кормов отмечены семена гречихи, донника, шренкии. Птенцы поедали также ягоды эфедры и дикой вишни, а из подземных частей растений в их пище отмечены только луковицы иксиолириона и из зеленых кормов — кусочки листьев травянистых растений.
У одного из 2-дневных птенцов в составе пищи обнаружены личинки саранчевых, мелкие частицы двукрылых и муравей. Зоб второго птенца был пуст, а в желудке обнаружены мелкие остатки семян растений и мелкие гастролиты. У 5-дневного птенца в зобу были только семена, а в желудке — их перетертые остатки. У 8-дневного птенца, кроме личинок саранчевых и двукрылых, обнаружены цикадка, коровка, златки, долгоносик и чернотелка, в желудке — остатки животных и растительных кормов. У 11-дневного птенца в пище встречены уже подземные части растений — мелкие луковицы иксиолириона, а кроме того,клопы и семена растений. Птенцы (5 особей) в возрасте 14—21 дня поедали ягоды эфедры и дикой вишни, а из животных — крупных жуков (жужелиц). В их зобах обнаружены также семена гречихи.
К концу первого месяца жизни состав кормов становится все более разнообразным: появляется зелень и различные моллюски, семена донника и липучки. Как и раньше, поедаются клопы, прямокрылые, жуки, ягоды эфедры и луковицы иксиолириона, но в большем количестве. Сердняя масса гастролитов в желудках таких птенцов составляет 0.5 г. В 1.5—2-месячном возрасте соотношение животных и растительных кормов в рационе кеклика постепенно выравнивается и в 3-месячном возрасте мало чем отличается от пищи взрослых особей.
На юго-востоке Казахстана выявлено небольшое, но устойчивое преобладание самцов (1:0.9) в различных популяциях: из 925 экз. 486 (52.6 %) оказались самцами. Такие же данные получены при вскрытии 36 однодневных птенцов из 3 гнезд (19 самцов, т.е. 52.8 %). Соотношение же возрастных групп зависит от сезона года. По материалам, полученным в весенние периоды 1974—1978 гг. в горах Чулак, на каждую наседку, успешно закончившую насиживание, в среднем приходилось от 11 до 14.7 птенцов. Однако такое соотношение довольно далеко от истины, поскольку эффективность гнездования у кеклика составляет 37.8-62 %, т.е. в среднем половина наседок теряет кладки. Расчеты показали, что на каждые 100 взрослых птиц, при равном соотношении полов, должно быть 66 насиживающих птиц — 50 самок и 16 самцов. Учитывая, что птенцы вылупливаются только из половины кладок, а на каждую наседку приходится в среднем 12.4 птенца, можно считать, что 100 взрослых птиц дают 400 молодых, т.е. соотношение взрослых к молодом птицам после периода размножения должно быть около 1:4, что и подтвержается на практике. Так, по данным осецних отстрелов, в Заилийском Алатау (ущелье Актерек) 13—14 ноября 1976 г. соотношение взрослых и сеголеток было равно 1:3.25 (51 особь), а в начале ноября 1978 г. в ущелье Жаманты соотношение это составило 1:3.45 (145 особей).
Календарные сроки разбавки зимних стай кекликов и образования пар во многом обусловлены высотой местообитания над уровнем моря и ходом весны. В стаях кекликов оживление наступает с первыми же теплыми днями. С восходом солнца громкие крики птиц раздадутся на склонах и на дне ущелий, часто птицы из одной стаи перелетают к другой, а иногда слышится писк самок, преследуемых самцами. Перезимовавшие стаи мельчают постепенно: сначала отмечаются мелкие стайки, потом пары. Первые встречи кекликов в горах Чулак отмечены в разные годы с 5 по 11 марта.
Мечение особей показало определенную привязанность отдельных птиц к определенной территории где они держались как зимой, так и в период размножения. Первое время-образовавшиеся пары мало заметны и не проявляют агрессивности по отношению друг к другу. Однако к мрменту появления яиц в гнездах между ними иногда вспыхивают ожесточенные драки (Рустамов, 1945; Попов, 1959; Остапенко, 1965). Схватки в брачный период имеют место только между самцами, занимающими одинаковые ступени иерархической лестницы. В Джунгарском Алатау у небольшого числа самцов (4.6 %) установлена полигамия, и было выяснено, что доминанты своим голосом подавляют активность и звуковую сигнализацию более слабых самцов и занимают наиболее удобные гнездовые участки.
Песня самца состоит из хорошо известного квохтанья и заканчивается «скирканьем» — «скрюююу», которое повторяется несколько раз подряд. Иногда в конце песни добавляются звуки «кро-о, кро-о», чем-то напоминающие голос вороны. У поющей птицы перья на шее топорщатся, туловище вытянуто вверх и немного вперед. Вокальная активность к полудню, чослабевает, и во второй половине дня, как и в дождливые и ветреные дни, песни слышатся редко.
На большей части ареала кеклики приступают к строительству гнезд в апреле. Гнезда, видимо, строят только самки. Как исключение, используются старые, более чем трехлетней давности постройки, лоток которых предварительно очищается от строительного материала. Гнезда устраиваются обычно на склонах, предпочтение отдается южной, юго-западной и юго-восточной экспозициям. Из 145 найденных гнезд 124 (85.6 %) располагались на склонах. 15 гнезд (10.3 %) были найдены в непосредственной близости от вершин и только 6 (4.2 %) располагались на дне ущелий.
Гнезда помещаются на открытых участках склонов у основания куста, под навесом камня, на уступе скалы, в траве. Одно гнездо было найдено даже в дупле старой арчи на высоте 1 м от земли (Сопыев, Караев, 1979). Зачастую гнезда находятся на значительном расстоянии от водных источников, иногда в нескольких километрах. Следует отметить, что в период строительства гнезд, откладки и насиживания кеклики довольно редко посещают водопои; поедаемые в это время зеленые части растений полностью удовлетворяют потребности птиц во влаге.
Появление первых яиц обычно совпадает с установлением положительных ночных температур. В горах Чулак первые яйца появлялись в гнездах в различные годы 3—18 апреля. В период откладки яиц самцы часто преследуют самок. Обычно после песни самец приближается к самке и начинает бегать вокруг нее, голова при этом низко опущена к земле и составляет прямую линию с вытянутой шеей. Крыло приспущено и касается земли. Брачные игры часто заканчиваются спариванием, продолжающимся 4—7 с. В некоторых случаях спаривание происходит без предварительного ухаживания самца. В это время большую часть суток птицы проводят в стороне от гнезд, чаще в 50—150 м; иногда в течение дня пары встречаются в одних и тех же местах. Приближаясь к гнезду, они проявляют величайшую осторожность и, заметив опасность, затаиваются или улетают.
Плотность расположения гнезд определяется весенней численностью птиц. Среднее расстояние между гнездами в горах Чулак составляло в разные годы 380—460 м. В Чу-Илийских горах гнезда с кладками располагались в 100—150 м друг от друга. На противоположных склонах расстояние между ними иногда сокращалось до 100 м и даже 10 м (Кузьмина, 1955).
Яйца бледновато-охристые с большим разнообразием оттенков. По основному фону разбросаны коричневые или темно-бурые пятна различного размера. Индивидуальная изменчивость в окраске яиц наблюдается даже в одной кладке. Со временем скорлупа выцветает. По наблюдениям за одной из брошенных кладок поверхность яиц сохраняла свою окраску в течение 20 дней и полностью обесцветилась за 50 дней. Форма яиц овальная. Размеры (п=186): длина 37—43.6, в среднем 39.93±0.08; ширина 27.6—31.7, в среднем 30.6±0.06. Масса свежих яиц (п=53) 18.7—21.6, в среднем 20.1±0.6. В процессе насиживания она уменьшается на 10.95 % и на 18—23 день равна 15.2—20.3, в среднем 17.9+0.27 т.е. 89.05 % от первоначальной массы. Средний интервал между откладкой яиц в гнезде составляет 32 ч (11 наблюдений). В 3 случаях яйца откладывались в гнезде разными самками (интервал между откладкой последующих яиц менее 24 ч), но впоследствии кладку насиживала только одна.
Величина полной кладки подвержена значительным изменениям и колеблется от 7 до 21 яйца, но средняя величина кладки в одной и той же местности меняется по годам незначительно. Так, средняя величина кладки в Казахстане составила 14.4 яйца, с колебаниями в отдельные годы от 12.6 до 14.9 (70 гнезд).
Птицы приступают к насиживанию к день откладки последнего яйца (13 гнезд из 18) или на следующий день (3 гнезда). В 2 гнёздах после начала инкубации добавилось по 2 яйца.
Насиживающие самки зачастую подпускают человека на расстояние до 1 м, а иногда не сходят с гнезда даже в тех случаях, когда до нихдотрагиваются. В жаркие часы дня у насиживающей птицы часто наблюдается учащенное дыхание, связанное с избытком тепла: клюв приоткрыт, горло заметно вибрирует. О режиме инкубации дает представление следующее наблюдение: самка за 11 часов насиживания отлучалась от гнезда на 13—15 % времени, на переворачивание яиц затрачивала 8—10 % времени, производя его каждые 40—50 мин (Киргизский Алатау, — Сагынбеков, 1976). За долгие часы насиживания у самок в прямой кишке скапливается большая масса экскрементов, выделяемых позднее на местах кормежек в виде массы, размерами и формой напоминающей яйцо — так называемый «гнездовой помет», первые находки которого помогают установить сроки начала насиживания.
Насиживание длится 23—25, в среднем 24 дня (11 наблюдений). Возможно, что на продолжительность инкубации влияет частота и продолжительность отлучек насиживающей птицы и погодные условия.
С момента появления первого птенца до вылупления последнего проходит 3—5 ч, а начиная с проклева 1-го яйца и до ухода выводка 1—2 сут (15 гнезд). Еще находясь в гнезде под наседкой, птенцы иногда покидают его на короткое время, удаляясь до 0.5 м.
Естественный отход яиц (неоплодотворенные, с погибшим эмбрионом) сравнительно невелик и составляет от 2.3 до 12 %, в среднем 6.2 %. В некоторых случаях наседки уводят выводки, оставляя в гнездах надклюнутые яйца с живыми, вполне сформировавшимися птенцами, отставшими в развитии всего на несколько часов.
По наблюдениям за 36 гнездами, из которых вывелись птенцы, только в 14 (38.8 %) выход составил 100 %, в остальных 22 (61.2 %) после ухода выводков оставались неоплодотворенные яйца (14 гнезд) и яйца с погибшими эмбрионами (7). Неразоренные гнезда составили 37.5 — 73.3 %, в среднем 57.5 %.
Гибель гнезд происходит в большинстве случаев в начале насиживания. Из 18 гнезд, подвергшихся разорению, только однажды яйцо исчезло на следующий день после появления его в гнезде. Во всех остальных случаях гибли полные кладки.
В году бывает только одна нормальная кладка, при гибели которой кеклики могут гнездиться вторично. Например, 31 мая в горах Чулак добыта самка с наседным пятном, у которой в яйцеводе было обнаружено яйцо (Кузьмина, 1955). Этот факт так же, как и встречи в природе поздних выводков, подтверждает возможность повторного размножения.
Двойное гнездование у птиц встрёчается исключительно редко. Оно установлено у 2 видов Alectoris: A. graeca и A. rufa (Portal, 1924). О кеклике до сих пор было известно только, что у выводков добывали самцов с наседными пятнами (Зверев, 1948; Кузьмина, 1955; Беме, 1975).
По нашим данным (Грачев, 1983), двойное гнездование свойственно и кеклику. Из 31 насиживавшей птицы, отловленной на гнездах в горах Чулак, 12 (38.7 %) оказались самцами. Кроме того, удалось установить принадлежность 8 гнезд 4 парам птиц. Наблюдения показали, что некоторые пары устраивают по 2 гнезда. Кладку в первом из них насиживает самец, во втором — самца. Возраст отловленных на гнездах самцов был около года, т. е. они впервые принимали участие в размножении. Иногда самцы приступают к насиживанию уже спустя 3—4 дня после откладки последнего яйца в первом гнезде, когда самка продолжает нестись во втором. Возможно, что при этом она спаривается с другим самцом и если это так, то здесь мы сталкиваемся с явлением частичной полиандрии.
Успешность гнездования у кеклика значительно колеблется по годам. В горах Чулак она колебалась в разные годы от 37.8 % (111 яиц в 9 гнездах в 1974 г.) до 62 % (245 яиц в 15 гнездах в 1977 г.).
Масса только что вылупившихся птенцов 11—14.65, в среднем 14.5 (п=56 экз.). Среди птенцов одного выводка разница в массе может достигать 2.9. Птенцы в этом возрасте имеют большой желточный мешок (до 13 % от массы тела) и развитую подкожную жировую клетчатку, которые сохраняются до 5-дневного возраста. В первые же 2 дня, а у некоторых только на третий, отпадает яйцевой зуб. В первые 2—3 дня жизни масса птенцов снижается в среднем на 1.2, но к концу первой недели уже достигает 20.8.
На 8-й день птенцы начинают подпрыгивать и пролетать до 50 см, на 13-й отмечено первое планирование над склоном на расстояние до 15 м. Выводок посещает водопой только с недельного возраста. Чем старше становятся птенцы, тем чаще они посещают водопои.
В большинстве случаев при выводке находится одна взрослая птица, но нередки случаи, когда их бывает и две (в среднем 26.4 % встреч выводков в Казахстане — от 9.5 до 29.8 % в разные годы). Можно предполагать, что такие выводки представляют собой объединение выводков при двойном гнездовании. Иногда в объединенных выводках можно встретить разновозрастных птенцов, а также группы из 4—6 взрослых птиц из числа не участвовавших в насиживании и временно присоединяющихся к выводкам.
При появлении опасности быстро убегают вверх по склону или улетают, в некоторых случаях затаиваются. Бегущие птицы легко преодолевают каменистые осыпи и участки отвесных скал. Вниз по склону бегают реже.
Полет используют, если появляется необходимость быстро достигнуть дна ущелья или уйти от внезапной опасности. Взлетая со склона, птица переходит на стремительное скольжение.Взлет начинается с частых взмахов, которые чередуются с полетом на неподвижно расставленных крыльях. Максимальное расстояние, которое могут пролететь кеклики, взлетевшие с вершины горы, составляет около 2 км (Попов, 1960).
По рыхлому и глубокому снегу передвигаются с трудом и в многоснежные зимы становятся легкой добычей различных хищников. В местах, где их преследуют, кеклики проявляют особую осторожность, но если их не беспокоить, они зачастую живут в непосредственном соседстве с человеком.
Кормятся на земле, употребляя в пищу в основном растительный корм и в меньшей степени беспозвоночных животных. В очень редких случаях могут кормиться на деревьях. Способы добычи корма разнообразны. Зеленые части растений и плоды сначала захватываются клювом, а затем отрываются. Расположенные в верхнем слое почвы подземные части растений птицы раскапывают попеременным движением лап. Находящиеся глубже в земле мелкие луковицы извлекают целиком, а крупные достают по частям, выдалбливая клювом в почве вертикальные ямки 8—10 см глубиной.
Большое значение в жизни кекликов имеют водопои, которыми они в основном пользуются в июле—сентябре. Частота посещений зависит от погодных условий и времени года. Весной, когда в пище преобладают зеленые части растений, птицы у воды встречаются редко, а в зимний период они обходятся без воды, склевывая снег.
Дневная жизнь кеклика зимой сводится к добыванию корма. Только в ясные, относительно теплые дни их можно видеть неподвижно сидящими на вершинах скал. Еще в предрассветные сумерки с мест ночевки слышны голоса кекликов. Перекличка занимает обычно несколько минут. С восходом солнца среди стаек наблюдается оживление и птицы начинают слетать к подножью склонов и на дно ущелий, где есть свободные от снега места. Здесь птицы проводят большую часть дня. К вечеру поднимаются к местам ночевок, расположенным под навесами скал или у кромки кустов.
Иногда в течение всей зимы стайка ночует на одном и том же месте, которое покрывается толстым слоем помета. Некоторые стайки в течение дня не покидают участков размером 200—300 м. Водопои зимой посещают редко, скорее случайно-, потребность в воде удовлетворяют, склевывая снег. Сильные снегопады пережидают в кустарниках или под навесами скал. В случаях длительного ненастья могут голодать по 2—3 дня. Перед снегопадами усиленно кормятся, пока снег не покроет оголенные участки земли. Особняком стоит интересное сообщение Р. Г. Пфеффера, который наблюдал зимой 1974/1975 гг. в Алмаатинском заповеднике за небольшой стайкой кекликов из 10—15 птиц, регулярно кормившихся в дневные часы у подножья склона среди высохших стеблей татарника. Здесь площадка 15X15 м под слоем снега была изрыта ходами, и иногда при подходе приходилось выпугивать птиц, взлетавших из-под снега. По мнению Р.Г. Пфеффера, кеклики делали в снегу ходы в поисках семян татарника.
К концу февраля, когда обнаженных от снега участков становится больше, кеклики перестают встречаться большими стаями (зимой в особо кормных местах концентрируется подчас по 100—150 птиц) и к началу марта начинают разбиваться на пары.
Сезонные перемещения. Наряду с известными вертикальными кочевками, имеющими место осенью и весной, в Таджикистане известны перемещения на многие десятки километров, которые можно рассматривать, как дальние перелеты (Попов, 1959). Они начинаются уже в первых числах сентября из глубины гор Кухистана и завершаются в декабре после достижения мест зимовок в низкогорьях южного Таджикистана.
Эти перемещения вызваны тем, что в горах Центрального Таджикистана образуется зимой глубокий снежный покров. Сумма атмосферных осадков в местах гнездования перелетных популяций кеклика (800—1400 мм в год) в 3—4 раза выше, чем в местах зимовок. Весеннее возвращение кекликов в Кухистан происходит в мае, но в связи с большой гибелью птиц во время перелета и на зимовках их обратное движение мало заметно. Такие перелетные по сути дела популяции кеклика в горах Центрального Таджикистана (Попов, 1959; Ковалев, Попов, 1980) являются пока единственными в своем роде и в других частях ареала неизвестны.
Уже у 2-дневных пуховых птенцов хорошо заметны пеньки 7 первостепенных маховых ювенального наряда, а к месячному возрасту оперение состоит уже из 3 нарядов — остатков пухового, развитого ювенального и первых перьев взрослого наряда (появляется пенек 9-го первостепенного махового, относящегося уже к взрослому наряду). На 6-й день жизни начинают развертываться опахала первых пяти первостепенных маховых, 2—10 второстепенные маховые представлены кисточками. Одновременно идет развертывание кисточек рулевых перьев, больших и средних кроющих перьев крыла. В течение первых 4 недель интенсивно растут маховые, рулевые и контурные перья на значительной части тела. К концу 4-й недели пуховой наряд сохраняется только на голове, брюхе и крестце — он сразу же сменяется перьями дефинитивного наряда, минуя стадию ювенального оперения.
У взрослых птиц четко выражена одна летне-осенняя линька, продолжительность которой составляет 4—4.5 мес. Сроки ее зависят от участия в насиживании. Особи, которые в нем не участвуют или потеряли кладки, объединяются в стайки и начинают линьку. Насиживавшие птицы начинают линьку только через 10—15 дней после вылупления птенцов.
На средней линии брюшка у них появляется широкая полоса пеньков, а спустя несколько дней они появляются на боках шеи, спине и груди.
Одновременно наступает линька маховых и рулевых. Смена первостепенных маховых идет в дистальном направлении от 1-го к 10-му. Второстепенные маховые начинают сменяться после того, как 1—4 первостепенные маховые уже достаточно отрастут. В линьке второстепенных маховых наблюдаются индивидуальные отклонения — смена перьев может начаться с любого из первых 4 второстепенных маховых. Так, среди 19 особей из Джунгарского Алатау начало смены второстепенных маховых с 1-го или же с 1-го и 2-го пера отмечено у 2 особей (10.5 %), со 2-го или со 2-го и 3-го — у 6 (31.5 %), с 3-го или 4-го — у 3 особей (15.8 %). У 8 птиц перья уже настолько отрасли, что установить порядок смены оказалось невозможным. У 5 особей (26.4 %) 2, 3 и 4-е оказались равной длины, а у 3 особей (15.8 %) наиболее длинными были 1, 2 и 3-е.
Линька проксимальных второстепенных маховых наступает несколько позже. Чаще она начинается с 10-го пера, но иногда с 9-го и идет, как правило, в 2 направлениях — дистальном и проксимальном. Отмечены случаи некоторой задержки линьки плечевых перьев. В целом же в линьке взрослых птиц остается еще много неясных моментов. В зимнее время и весной встречаются особи, у которых в области шеи и на спине остаются отдельные пеньки и кисточки (Дементьев, 1952; Карташев, 1952 г; Кузьмина, 1955). В Чу-Илийских горах в феврале из 50 осмотренных экземпляров линька отмечена у 5, а в апреле у 28 птиц из 40 (70 %), причем характер этой линьки остается еще не выясненным (Кузьмина, 1955).
Подробнее о сокращениях можно прочесть здесь.
Литература
• «Линька воробьиных птиц Северо-Запада СССР». Рымкевич Т.А., Савинич И.Б., Носков Г.А. и др.; Изд-во Ленингр. ун-та. 1990 г.
Сайты
www.featherbase.info
Сайты
www.pbase.com, ibc.lynxeds.com, orientalbirdimages.org, www.fotoparus.com
Песня
…
Позывка
…
Электроманок
…
Сайты
www.xeno-canto.org
Perdix Graeca. Meisner. Syst. Verg. Vög., welche die Schweiz bewohnen etc., 1804, стр. 41, ex Daubenton PI. Enul., 231, Греция.
Синонимы.
Caccabis saxatilis. Meyer. Naturg. Vog. Deutschl., 1, 1805—1809, стр. 86.
Tetrau kakelik. Falk. Beytz. z. topogr. Kenntn. Russ. Reichs, III, 1786, стр. 390 (nomen nudum).
Perdix Chukar. Gray. 111. Ind ZooL, 1, 1832, табл. 54.
Русские названия. Каменная куропатка — название книжное. Кеклик — широко распространенное у народов Средней Азии звукоподражательное название этого вида, в конце XVIII в. перешедшее в русский язык.
Ареал. Южная Европа, Передняя, Средняя и Центральная Азия: от Пиренеев и Альп, Сицилии, Балканского п-ва и островов Средиземного моря через Малую Азию, Кавказ, Копет-Даг, Гиндукуш, Памир, Тянь-Шань и Алтай до Маньчжурии, Тибета и Гималаев.
Характер пребывания. Оседлая птица. При выпадении снега в горах предпринимает незначительные вертикальные кочевки, переходя на малоснежные участки.
Биотоп. Глубокие ущелья и каменистые склоны гор от верхней границы леса до вечных снегов, каньоны рек в горах и предгорьях; по каменистым россыпям, выходам скал и вертикально расчлененным участкам рельефа проникает в предгорья и на прилежащие равнины, по скалистым останцам и глинистым холмам заходит и в песчаные пустыни.
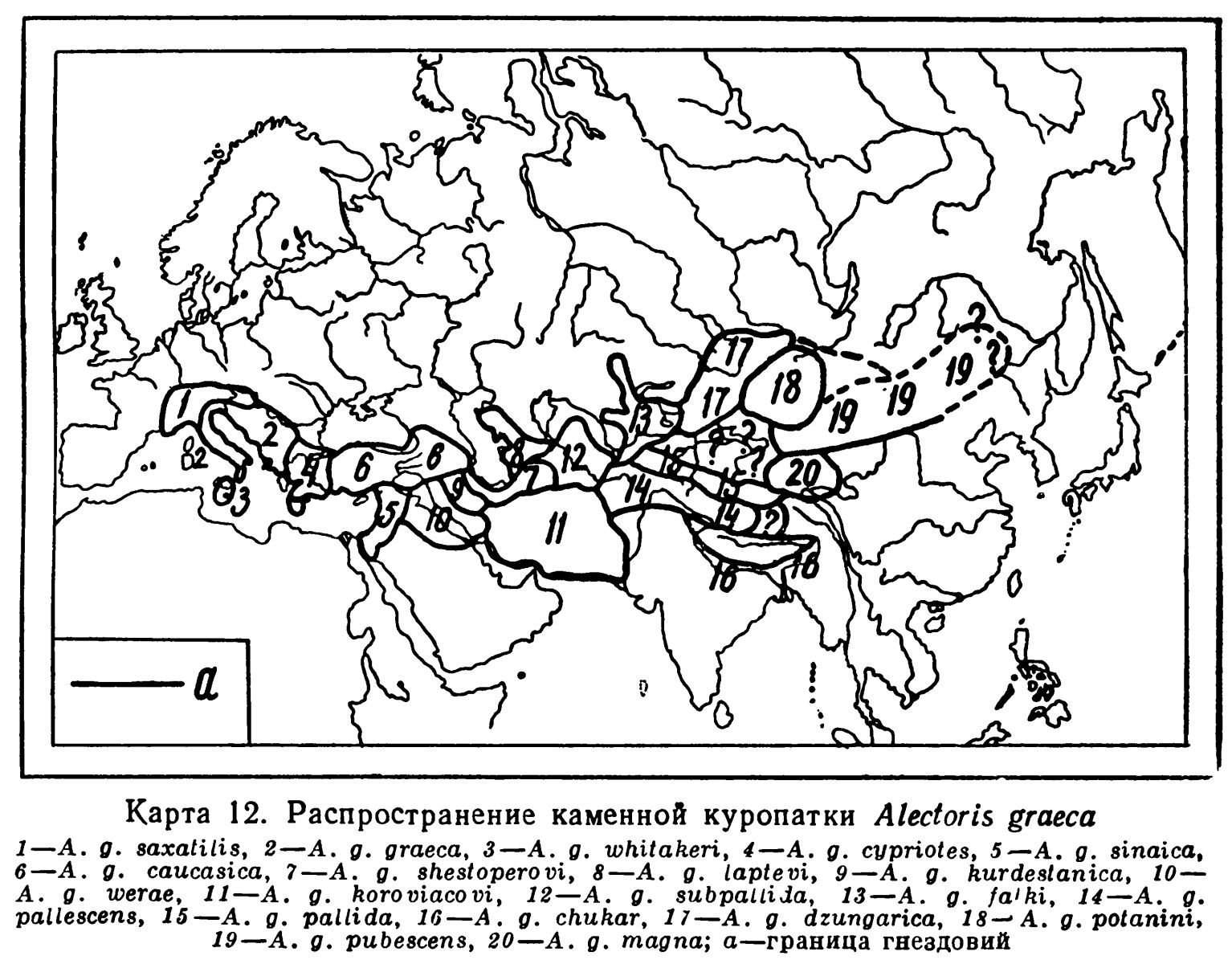
Подвиды и варьирующие признаки.
— По всем Альпам на восток до Юго-Восточной Венгрии распространена A.g. saxatilis Meyer, 1805—1809;
— от Апеннинских гор (Италия) до Югославии, Греции, Албании, Западной Болгарии и Ионических о-вов распространена A.g. graeca Meisner, 1804;
— в Сицилии A.g. whitaneri Sсhiebel. 1934;
— на о-вах Эгейского моря, на о-ве Кипр, в европейской части Турции и Юго-ВосточнойБолгарии, в западных и южных частях Малой Азии, включая Тавр, A.g. cypriotes Нагteгt, 1917;
— от Синайского п-ва до Сирийского побережья A.g. sinaica Bonaparte, 1856,
— Среднюю и Восточную Малую Азию, Кавказ и Закавказье населяет A.g. caucasica Suschkin, 1927;
— по горам южного берега Каспийского моря и в Копет-Даге живет A.g. shestoperovi Susсhkin, 1927;
— западную часть Туркмении, Большие и Малые Балханы, Мангышлак и Западный Узбой заселяет A.g. laptevi Dementiew, 1945;
— в Южном Курдистане A.g. kurdestanica Meinertzhagen, 1923;
— в Юго-Западном Иране и прилегающих районах Месопотамии A.g. werae Sarudny et Loudon, 1904;
— в Восточном Иране, включая Паропамиз и его отроги, в Белуджистане и в Синде A.g. koroviakovi Sarudny, 1914;
— в Кызыл-Кумах, Кугитанге и Баба-Таге A.g. subpallida Sarudny, 1914;
— горы системы Тянь-Шаня населены A.g. falki Нагteгt, 1917;
— в Гиндукуше от Гильгита, через Балтистан до Ладака A.g. pallescens Hume, 1873;
— в Кашгарии и возможно в пограничных участках Памира и Семиречья, в Западном Куэньлуне, Алтын-Таге и Западном Нань-Шане A.g. pallida Hume, 1873;
— в Гималаях и Непале A.g. chukar Gray, 1830—1832;
— Северное Семиречье, Тарбагатай, Алтай, Западный Саян, Танну-Ола и прилегающая Монголия населены A.g. dzungarica Susсhkin. 1927;
— от Алашаня до Монгольского Алтая и Хангая распространен A.g. potanini Suschkin, 1927;
— в Гань-су и прилегающих районах Китая вплоть до Маньчжурии A.g. pubescens Swinhое, 1871;
— в Цайдаме, Куку-норе, Южно-Куку норском хребте, Бурхан-Будда и в истоках р. Хуанхэ A.g. magna, Przewalski 1876.
Отличия подвидов сводятся к удовлетворительно или плохо заметным вариациям особенностей окраски и к небольшим различиям в размерах. Географические изменения экологии (календарь периодических явлений, трофические связи и т.п.) известны недостаточно.
Литература
• «Птицы Советского Союза». Дементьев Г.П., Гладков Н.А., Исаков Ю.А., Карташев Н.Н., Кириков С.В., Михеев А.В., Птушенко Е.С. Под общей редакцией Г.П. Дементьева и Н.А. Гладкова. Изд. «Советская наука» Москва. 1952 г. том 4. стр. 154-173.
Литература
• Коблик Е.А. 2025. Фауна птиц стран Северной Евразии в границах бывшего СССР: Списки видов.
• IOC World Bird List v15.1 (Updated February 20, 2025) by Frank Gill, David Donsker & Pamela Rasmussen.
• Н.Н. Балацкий. Птицы Северной Евразии. Таксономический список: август 2023 г.
• «Птицы СССР. Курообразные. Журавлеобразные». т.2. Р.Л. Беме, Н.П. Грачев, Ю.А. Исаков, А.И. Кошелев, E.Н. Курочкин, Р.Л. Потапов, А.К. Рустамов, В.Е. Флинт. Ленинград. Издательство «Наука». 1987 г. стр. 70-86.
Сайты
• zmmu.msu.ru
• www.balatsky.ru
• www.worldbirdnames.org
• Avibase by Denis Lepage
• Zoonomen by Alan Peterson
• Taxonomy by John Boyd
• сайт Василия Вишневского
• www.xeno-canto.org
• www.featherbase.info, www.pbase.com, www.macaulaylibrary.org