
Глухарь — самая крупная из куриных птиц СССР, ростом почти с индюка. С земли поднимается тяжело, громко хлопая крыльями. Летит обычно над самым лесом или в полдерева; лишь осенью совершая более значительные перемещения, летит высоко над лесом.
Семейство: Тетеревиные Tetraonidae
Род: Глухари Tetrao
Вид: Tetrao urogallus (Linnaeus, C 1758)
Протоним: Tetrao Urogallus.
Синоним: Tetrao major, Tetrao urogallus, Tetrao urogallus major, Tetrao urogallus urogallus.
Немецкое название: Auerhuhn
Испанское название: Urogallo Común.
Казахское название: меңіреу құр, меңіреуқұр, Мењіреу құр, саңырау құр
Украинское название: Глушець.
Другие названия: обыкновенный глухарь, белоклювый глухарь, глушец, глушак, глушень, глухой тетерев, моховик, глухарка, глушица, копалуха, курица, мошняк.
Taxonomic Serial Number : 553893 | Global Biodiversity Information Facility : 2473577.
Список ПОДВИДОВ вида Глухарь Tetrao urogallus (Linnaeus, C 1758)
‐ Поволжский глухарь — Tetrao urogallus volgensis (Buturlin, SA 1907)
‐ Скандинавский глухарь глухарь — Tetrao urogallus urogallus (Linnaeus, C 1758)
‐ Светлоклювый глухарь — Tetrao urogallus kureikensis (Buturlin, SA 1927)
‐ Южноуральский глухарь — Tetrao urogallus uralensis (Nazarov, PS 1886)
‐ Сибирский глухарь — Tetrao urogallus taczanowskii (Stejneger, LH 1885)
ОСНОВНОЕ ОПИСАНИЕ ПО: Глухарь — Tetrao urogallus (Linnaeus, C 1758)
В зависимости от ширины светлых полосок в струйчатом рисунке общая окраска верха кажется то светлее, то темнее. Поскольку с возрастом струйчатость становится тоньше, самец становится более тёмным. Первостепенные маховые коричневые, с более светлым наружным опахалом, которое при изнашивании может стать совсем белым (особенно на 5–8 маховых). У второстепенных маховых перьев на внешних опахалах преобладает коричнево-серый струйчатый рисунок по коричневому фону, на самых внутренних второстепенных маховых этот рисунок есть и на внутренних опахалах; здесь же развит белый крап, образующий иногда беловатые поля.
Первое, иногда и 2-ое второстепенные маховые имеют у основания опахал 1–2 белых пятна, особенно часто у годовалых птиц. Малые, средние и большие кроющие крыла коричневые с поперечным струйчатым рисунком. Нередко вершины этих перьев имеют сероватый и даже белый цвет. Подмышечные перья белые, на самых длинных из них — чёрные вершинные пятна. Рулевые перья чёрные с белыми пятнами и волнистыми полосками в средней части пера, лучше развитыми на внешних опахалах (нередко отсутствуют у старых птиц на 1–3 центральных парах). Нередко центральная пара несёт, за исключением вершины, коричневый крапчатый рисунок, который у старых птиц менее заметен и может вообще отсутствовать. Верхние кроющие хвоста окрашены под цвет поясницы, со светло-серым струйчатым рисунком по коричневому фону и с вершинными белыми каймами шириной в 3–5 мм. Нижние кроющие хвоста чёрные с белыми вершинами, нередко испятнанными чёрным цветом или разбиты стержневой чёрной полосой на 2 вершинных пятна.
У старых самцов нередко эти перья сплошь чёрные. На нижней стороне тела выделяется грудной пластрон зелёного цвета, с металлическим отливом. Этот пластрон образуют зелёные вершины чёрно-бурых грудных перьев. Передняя часть и середина брюха покрыты у старых птиц чёрными перьями с зеленоватыми блестящими каймами, а у более молодых птиц здесь развит серый струйчатый рисунок. Задняя часть брюха до подхвостья бурая, с сероватыми вершинами перьев. Посередине брюха часто развиты белые пестрины, которых иногда бывает настолько много, что они создают почти сплошную белую окраску. Бока коричневые с сильно развитым светлым струйчатым рисунком. Клюв беловатого или желтоватого цвета, пальцы серые, радужина коричневая.
Описанный наряд одевается после второй полной осенней линьки, но отдельные изменения в нем происходят и далее, сводясь в основном к потемнению окраски, утоньшению струйчатого рисунка и исчезновению или уменьшению в размерах ряда белых пятен.
Самец в первом осеннем наряде. Серый и белый цвета развиты больше, чем в последующих нарядах. Беловатый струйчатый рисунок несут перья «бороды», ряд перьев на груди, ограничивающих зелёный пластрон, центральная пара рулевых перьев, верхние кроющие хвоста. На брюхе много белых пятен, расположенных большей частью на внешних опахалах перьев. На рулевых перьях есть концевые белые полосы, особенно хорошо выраженные на 2–3 центральных и 1 наружной парах рулевых. На нижних кроющих хвоста чистые белые вершины. На внешнем опахале самого длинного пера крылышка мелкий крапчатый рисунок. На кроющих перьях крыла струйчатый серый рисунок занимает почти все вершины. Подмышечные перья чисто белые.
Взрослая самка. Верх пёстрый, из сочетания чёрных, желтовато-рыжих белесых и белых полос и пятен. Большинство перьев имеет поперечно-полосатый рисунок, с преобладанием струйчатого серого рисунка на вершине. Первостепенные маховые коричневые, с более светлым наружным опахалом, на котором имеется неясный поперечный рисунок. Второстепенные маховые перья с широкими белыми вершинными каймами, особенно широкими на самых внутренних перьях, и с охристым поперечным рисунком по коричневому фону. Большие, средние и малые кроющие крыла с белыми вершинами, особенно хорошо выраженными на проксимальной части крыла. Нижняя часть тела также пёстрая, подбородок беловатый, горло желтовато-рыжее, как и верхняя часть шеи. Остальная часть шеи пёстрая. На груди желтовато-рыжий пластрон, в нижней части которого есть чёрные поперечные полоски. Нередко такие полосы разбросаны по всему пластрону. Далее к брюху рыжий цвет пластрона переходит в пёструю окраску брюха, где доминируют широкие белые вершины перьев, благодаря которым здесь нередко преобладает белый цвет.
Бока тела окрашены так же, но вершины перьев нередко испятнаны чёрным. Рулевые перья с белыми вершинными каймами и поперечным рисунком из широких охристо-рыжих и чёрных полос. Верхние кроющие хвоста окрашены так же, нижние — несколько бледнее, с более широкими охристыми полосами. Клюв и пальцы тёмно-роговые. Самки имеют 2 типа окраски со всеми вариантами переходов между ними — рыжий и серый (эти типы нередко называют фазами). У «рыжих» преобладает желтовато-охристая окраска, грудной пластрон чистого ржаво-охристого цвета, без тёмных полос. Белые каймы и вершины перьев по всему телу выражены нерезко и иногда имеют желтоватый оттенок, тёмные полосы обычно бурого цвета. Поперечные полосы на нижней стороне тела крупные, оперение ног светлое, с желтоватым оттенком. Самки серого типа окрашены темнее, с более контрастной окраской белого и чёрного цветов. Грудной пластрон испещрён поперечными полосами чёрного и белого цветов, окраска ног тёмная с мелким рисунком. Эти 2 типа окраски встречаются в любой популяции, но в холодных континентальных областях преобладает серый тип, в более тёплых и влажных — рыжий. От возраста тип окраски не зависит.
Ювенальный наряд. Самцы. Верх головы тёмно-серый, с рыжеватым оттенком над бровями. Бока головы буроватые, с белым каплевидным рисунком. Кроющие уха серые. Горло грязновато-белое с тонким чешуйчатым рисунком по бокам. На спине преобладает буровато-охристый цвет, здесь развит поперечно-полосатый рисунок из тёмно-бурых и беловато-охристых полос. Нижняя часть спины, поясница и надхвостье более серые и по центру испятнаны белым: на перьях здесь развиты характерные наствольные полоски, расширяющиеся к вершине. На груди рыжевато-бурый пластрон с более тёмной буроватой серединой. Брюхо грязно-серого цвета с неясными бурыми пятнами.
Бока светлые или тёмно-бурые, с рыжеватой поперечной полосатостью и концевыми белыми пятнами на перьях. Первостепенные маховые тёмно-бурые с поперечным рисунком из коричневых полос на внешнем опахале и на верхней части внутреннего. На второстепенных маховых поперечный рисунок более чёткий, образован желтоватыми полосами. На проксимальных второстепенных маховых рисунок более сложный, с краевыми волнистыми темно-бурыми пятнами и вершинным белым пятном. Большие, средние и малые кроющие крыла охристо-коричневые, с неясным поперечным рисунком или лёгкой буроватой струйчатостью, и с наствольной белой полоской, расширяющейся к вершине.
На проксимальных партиях этих перьев рисунок также усложнён бурыми пятнами. Рулевые перья в верхней части поперечно-полосатые, с белыми вершинными каймами. В нижней части пера этот рисунок едва намечен. Молодые самки отличаются прежде всего большим развитием рыжих тонов в оперении и грудным пластроном чистого желтовато-рыжего цвета. Рулевые перья несут поперечный рисунок почти по всей длине. На нижних кроющих перьях первостепенных маховых хорошо развит поперечно-полосатый рисунок, а на самых проксимальных перьях развиты белые поля, тянущиеся вдоль стержня и внутреннего края. В окраске маховых перьев, несмотря на указания Хёглунда (Hoglund, 1952), отличий от окраски самцовых перьев не обнаружено.
Пуховой птенец. На верхней стороне тела довольно сложный узор из черноватых и охристых полос, среди которых особенно выделяется охристая полоса с черноватым окаймлением поперёк верхней части спины, охристая полоска на пухе, кроющем крыло, и тёмно-охристая полоса с бурым окаймлением, идущая по спине. Узор на голове состоит из чёрных пятен у основания клюва, над бровями, у глаза и около ушного отверстия, неясных чёрно-бурых штрихов на темени, нескольких полосок на затылке и V-образной полосы на лбу, направленной вершиной назад. Нижняя часть тела желтовато-охристая, с неясной охристой перевязью на груди и охристым налётом на подхвостье. Суточная активность, поведение. В зимнее время большая часть дня (до 22,5 часов) проводится под снегом, кормёжек либо одна в середине дня, либо две — утром и вечером. В период токования птицы особенно активны на вечерних и утренних зорях. Покидая токовище около 9 часов утра, птицы кормятся 2–3 часа, затем не менее 3 часов отдыхают и потом вновь кормятся до вечера. В летнее время суточная активность изучена недостаточно, но в целом она имеет сходную ритмику с пиками кормовой активности на рассвете и в предвечерние часы.
Враги, неблагоприятные факторы. Ни один из хищников существенно не влияет на численность глухаря. Взрослые птицы терпят незначительный урон только от крупных хищников: куницы и соболя, лисицы, волка, росомахи, филина, беркута и орлана-белохвоста. Молодые птицы, особенно в возрасте 2–3 месяцев,становятся жертвами значительно большего количества хищников, среди которых на первом месте стоят ястреб-тетеревятник, сарыч, куница, соболь и горностай. В ряде мест на западе Европы в связи с резко возросшей численностью кабана в охотничьих угодьях численность глухарей стала катастрофически снижаться — кабаны стали уничтожать значительное число кладок.
Из неблагоприятных факторов на первом месте стоят сильные похолодания с длительными дождями во время массового появления птенцов. Не имея ещё постоянной температуры тела, они всё время нуждаются в регулярном обогреве. Чем холоднее и дождливее погода, тем чаще они должны обогреваться под самкой, тем короче время на кормёжку и тем меньше доступен их основной корм — мелкие насекомые (Hoglund, 1955). Такие похолодания, нередкие на севере ареала («возвраты холодов»), в случае их большой длительности могут привести к гибели почти всех птенцов. Например, на Урале наибольшая корреляция численности глухарей со среднедекадными атмосферными температурами падает на вторую декаду июня (коэффициент корреляции + 0,534±0,036), то есть как раз на время вылупления и первых дней жизни птенцов (Данилов, 1975). На юге ареала заметный ущерб численности глухарей может быть нанесён засухами.
Хозяйственное значение, охрана. Практическое значение глухаря для человека в настоящее время заключается именно в той роли, которую он играет в экосистемах бореальных лесов. Кроме того, он до сих пор сохранил в отдельных районах промысловое значение, хотя и очень ограниченное. Так, по неполным анкетным данным, в 1960 году на 70% территории РСФСР было добыто 300 000 глухарей, что, по-видимому, составляет менее половины действительного числа добытых птиц этого вида на всей территории РСФСР.
Тем не менее и в прежние времена заготовки глухарей, в том числе и на экспорт, были во много раз ниже других тетеревиных птиц, прежде всего благодаря худшему качеству мяса. В настоящее время развитие промысловых заготовок глухаря нерентабельно. В то же время постоянно возрастает роль этого вида как объекта спортивной охоты. Оседлость, широкая естественная кормовая база и та высокая численность, которой может достигать глухарь в благоприятных условиях, делает этот вид одним из наиболее перспективных для охотхозяйств лесной зоны СССР. Главные биотехнические мероприятия заключаются прежде всего в сооружении специальных подкормочных площадок, где ведётся регулярная подкормка в зимнее время (Борискин, 1974) и, возможно, в создании специальных убежищ для молодых выводков на случай затяжных дождей. В организации охоты необходимо существенное ограничение (если не полное исключение) весенней охоты на токах, которая может допускаться только после того, как самки перестают посещать токовище. При такой практике совершенно необходима резервация определённых токовищ, куда прекращается доступ человека на всё время токования.
Подпуск в угодья привозных глухарей следует считать крайне нежелательным. В крайних случаях его организация должна осуществляться по рекомендациям и под контролем опытных специалистов.
Реинтродукция глухаря в пределах СССР была успешно осуществлена только в одном случае, в борах у села Боровое, Северный Казахстан (Черепанов, 1981).
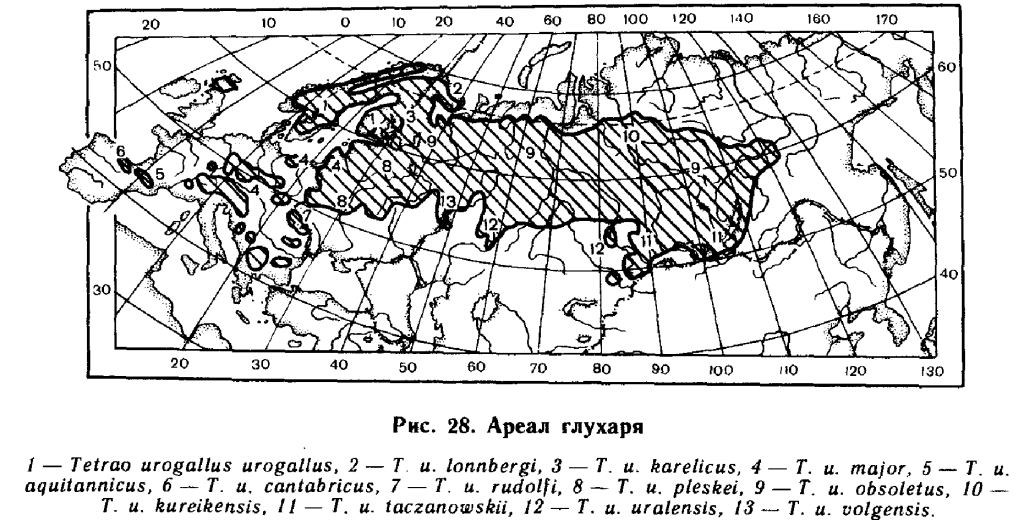
В Скандинавии она поднимается до 70° северной широты, на Кольском полуострове по крайней мере до 69°30′. От Кольского залива граница идёт на юго восток, включая в ареал леса в центральной части полуострова (бассейн озёр Имандра, Нотозеро и рек Туломы, Колмы, Умбы, окрестности Ловозера). По реке Поной встречается вниз по течению до посёлка Канёвки, есть у села Пялиць. К востоку от Белого моря граница начинается примерно на широте 65°, поднимаясь в бассейне реки Печоры выше Полярного круга (долина реки Шапкина). По западному склону Урала идёт на север по крайней мере до 66°30′ северной широты, встречен на реке Усе (Кириков, 1975). На восточном склоне встречается до станции Красный Камень (Данилов, 1975). Далее к востоку известен под Салехардом; на севере Красноярского края поднимается до южных районов лесотундры, регулярно гнездясь до 67°30′, а по долине Енисея — до 71°30′ (Яковлев, 1930). По последним данным, глухарь по реке Таз достигает только Полярного круга, а по Енисею — устья Хантайки, 68° северной широты (Назаров, 1968; Сыроечковский, Рогачёва, 1968). Восточнее Енисея северная граница не выяснена, видимо, она спускается к югу к долине реки Нижняя Тунгузка, севернее пока не встречен.
Дальше к востоку глухарь встречается по реке Вилюй, особенно в верховьях и по притокам Орт-Юрях, Ньохчук, Кемпендяй, Чона и по водоразделу между Леной и Вилюем в урочищах Намана, Оргольой, Быныттах (Андреев, 1953). Восточнее города Вилюйска не найден. Отсюда граница идёт на юг, к реке Лене, где этот вид найден в долинах рек Чара и Токко, и далее к северо-восточному берегу Байкала, где он есть всюду в прибрежной части озера. Населяет лесной пояс Хамар-Дабана. Наиболее восточная точка в Забайкалье — устье реки Муи (Поляков, 1873). От южного Прибайкалья граница идёт к Кяхте, к северному берегу озера Хубсугул и охватывая с юга истоки Енисея, идёт на запад по хребту Танну-Ола, где есть на северных склонах. Граница охватывает с юга горные леса Алтая (в Монгольском Алтае не найден), и идёт на север по реке Оби. Изолированный участок ареала — в Калбинском Алтае (Кузьмина, 1962). От долины Оби, под 56° северной широты граница идёт на запад, огибая с севера Барабинскую степь. По правобережью Иртыша в начале XX века водился в борах районов Семипалатинска (Белокаменск) и Павлодара (Хахлов, Селевин, 1928; Селевин, 1929; Залесский, Залесский, 1931). Ещё недавно он встречался в борах Курганской и Челябинской областей (Санарский бор у Троицка), населял кустанайские (Полферов, 1896) и кокчетавские боры (Штегман, 1934).
На южном Урале самая южная точка 51°30′, массив Шайтан-Тау (Кириков, 1952). Отсюда на запад граница идёт через Бузулукские боры к Жигулям, южнее которых глухарь ещё недавно водился в Черкасском лесхозе, 52°30′ северной широты (Болгов, 1968). Дальше граница поднимается к северу, к лесам Мордовской АССР. В Рязанской области есть только в северной части (Хомяков, 1901), сейчас в основном только в пределах Окско-террасного заповедника (Киселёв, 1971). В Калужской области в начале века глухарь обитал в Козельском, Жиздринском, Мосальском и Перемышльском районах (Филатов, 1915). В ареал входят также леса по левобережью Десны, в Карачевском и Трубчевском районах Брянской области (Граве, 1933; Суслова, 1937), и в самых северных районах Украинской ССР. На Западной Украине глухарь обитает в Карпатах, причём горные популяции изолированы от равнинных.
В Европе, вне пределов СССР, глухарь обитает в Польше, в северных, южных и юго-западных воеводствах: Белостокском, Ольштинском, Козалинском, Щецинском, Будгощском, Катовицком, Краковском, Вроцлавском, Жещувском, Гданьском и Люблинском (Marchlewski, 1962), в восточных частях ГДР, в лесах на ряде возвышенностей Западной Европы (Арденны, Рейнские Сланцевые горы, Гарц, Тюрингский Лес, Рудные горы, Судеты, Шумава, Франковская и Швабская Юра, Шварцвальд, Вогезы), в горных лесах Кантабрии, Пиренеев, Альп, Карпат, Родоп и Динарского нагорья. Обитал прежде, но был истреблён на Британских островах (в Англии — в XVII, в Шотландии и Ирландии — в XVIII веках), в Оверни (Франция) в XVIII веке (Andrew, 1958). В Шотландии был интродуцирован в 1837 году, и сейчас населяет большую часть её от Дорнох-Фирт до долины Форт-Клайд (Ваппегшап, 1963).
Численность. Данные табл. 5 показывают, что наиболее обычной является плотность в 10–16 птиц на 1 000 га. Другие данные о численности, превышающей указанную в 10–20 раз, видимо, показывают ту максимальную плотность, которую может достигать вид в благоприятных условиях обитания и при отсутствии воздействия антропогенного фактора. Следует учитывать, что даже в обширных массивах сосновых лесов глухари никогда не распределяются равномерно по всей площади, и места с их обилием чередуются с участками, где глухарей очень мало или нет совсем. Поэтому высокая численность птиц, до 20–25 на 100 га, может создаваться осенью или зимой только в отдельных местах их концентрации — на богатых ягодниках или на участках зимовки отдельных стай. Пересчёт же такой численности на более обширные области приводит к сильному завышению данных по численности этого вида.
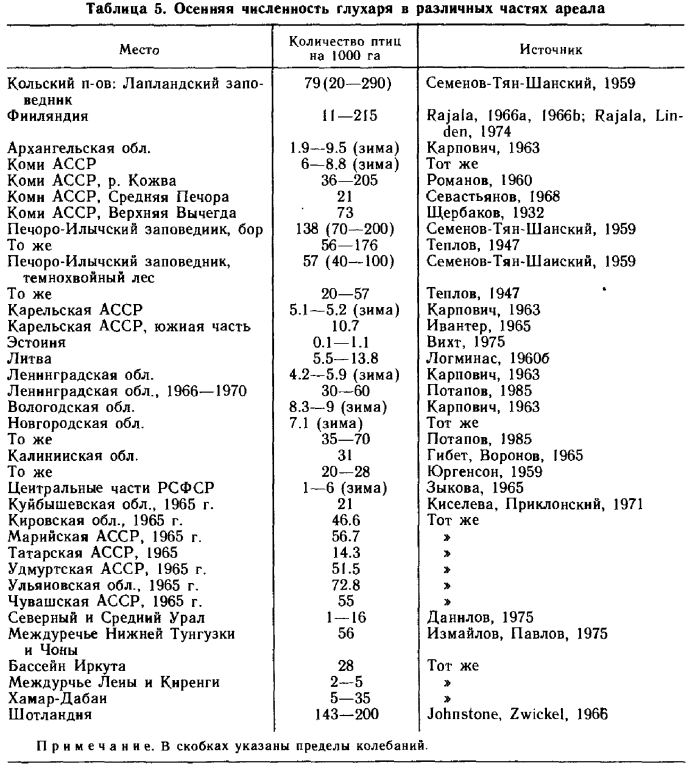
Колебания численности по годам происходят в различных частях ареала далеко не синхронно и с разной амплитудой. В целом, по южной половине ареала численность более стабильна, чем по северной. Наиболее оптимальной в отношении климатических условий для глухаря является подзона южной тайги в местах с явно выраженным континентальным климатом — в этих районах численность глухаря отличается наибольшей стабильностью.
Соотношение полов в популяциях близко к 1 : 1 (в Печоро-Илычском заповеднике 58 % самцов, — Теплов, 1947). По ряду данных, среди птенцов преобладают самки, но смертность их выше, чем самцов, и в старших возрастных категориях начинают преобладать самцы. Так, в Карелии среди молодых птиц самки составляют 57,3 %, среди взрослых 44,8 % (Ивантер, 1974).
Доля молодых птиц в популяции подвержена сильным колебаниям на севере ареала (от 3 до 81 %), тогда как в подзоне южной тайги возрастной состав более стабилен и молодые птицы составляют 53–80% (Семёнов-Тян-Шанский, 1959; Романов, 1975). В Финляндии например, за 4 года (1967–1970) процент молодых птиц осенью колебался от 45,8 до 62,5 (Pulliainen, Loisa, 1972). В Приокско-Террасном заповеднике в 1961-1966 годах процент молодых птиц изменялся в пределах 15,6–71,8 (Киселёв, 1971).
В пределах ареала населяет самые разнообразные типы леса, различной густоты, ярусности, увлажнённости, возраста основных пород, но в целом по ареалу отчётливо прослеживается связь с сосной, особенно в зимнее время. Явно избегает сплошной темнохвойной тайги, оказывая предпочтение сосновым борам или лесам с разнообразной экологической обстановкой — сочетаниям различных по возрасту участков леса, полян, гарей, болот (в особенности верховых). Сезонная смена мест обитания проявляется в основном перемещением на лето из сосняков в ельники. Характер рельефа не имеет существенного значения.
Глухари живут и на обширных, нередко заболоченных низменностях, особенно при сочетании последних с возвышенными грядами типа карельских сельг или озов, и в горной местности, причём в горах они поднимаются до самой верхней границы хвойного леса. По северной окраине ареала глухари выходят в разреженные лесные фитоценозы южных окраин лесотундры, но как правило, откочёвывают отсюда в зимнее время.
Весенние, летние и осенние корма гораздо разнообразнее и зависят в основном от растительности данной местности. Среди наиболее важных кормовых объектов — черника, Vaccinium myrtillus (стебли — осенью и весной, ягоды — летом и осенью), голубика, Vaccinium uliginosum (листья и ягоды — летом и осенью), пушица влагалищная, Eriophorum vaginatum (листья и цветы — весной и летом), сон-трава, Pulsatilla patens (соцветия — весной).
Весной глухари прежде всего поедают свежую зелень, цветы и соцветия трав, кустарников и деревьев. Характерным компонентом летнего рациона являются семена травянистых растений (осоки, марьянник, живородящая гречиха), листья и стебли хвощей и папоротников, а также листья различных ив, осины, берёзы, ольхи. Летом и в начале осени поедаются и животные: слизни, пауки, различные насекомые, среди которых предпочитаются кузнечики, муравьи и личинки пилильщиков, иногда даже ящерицы. В целом же доля животных кормов в питании глухаря невелика и не превышает в июле — августе 10%. Особенно охотно животную пищу потребляют линяющие птицы. Осенью основным кормом становятся ягоды.
Помимо них в ряде районов большое значение приобретает хвоя лиственницы, листья осины, кедровые орехи и жёлуди, а местами и злаковые культуры. Запасание гальки осенью для глухарей очень важно, и потребность в последней может явиться даже причиной массовых перемещений птиц, особенно в низинных районах, бедных обнажениями. Максимальное количество гальки в желудке самца 71 г, самки 27 г. Наиболее интенсивное собирание гальки начинается со второй половины сентября, в особенности после первых заморозков, когда глухари начинают переходить на зимний корм, и продолжается вплоть до установления снежного покрова. На утренних зорях птицы вылетают в поисках гальки на берега рек и ручьёв, на грунтовые дороги и карьеры, на обрывы террас и различного рода обнажения почвы. В течение зимы количество гастролитов в желудках постепенно уменьшается.
В питании птенцов в первые две недели жизни преобладают различные насекомые и пауки. Из растительных кормов первыми начинают поедаться цветы черники (в первую же неделю жизни), а начиная со второй недели, доля растительного корма увеличивается с каждым днем. В августе питание молодых птиц мало чем отличается от взрослых, в основном несколько большим количеством насекомых и зелени.
Самцы начинают расхаживать по насту в токовой позе, слегка отставив и опустив частично раскрытые крылья, которые чертят по снегу вдоль следов характерные борозды; иногда они прыгают и даже совершают токовые взлёты. Это возбуждение возникает спонтанно: если в группе кормящихся самцов начинает вдруг «чертить» по снегу и взлетать один самец, следом за ним то же самое начинают делать и другие. Такое начальное токование не привязано определённо к какому-нибудь месту, оно возникает и на местах кормёжек, и в местах ночёвок, когда утром птицы покидают свои лунки, но только очень редко происходит на самом токовище. Брачное оживление в таких ситуациях продолжается недолго, считанные минуты, но после него место, где «токовали» несколько самцов, напоминает по следам токовище в разгар токования. В случае присутствия самок, последние никак не реагируют на токовое поведение самцов.
Постепенно с усилением токовой активности самцы начинают всё чаще навещать токовище, сначала в утренние часы после восхода солнца, затем во всё более раннее время и, наконец, начинают посещать его регулярно, прибывая сюда вечером и ночуя на своём токовом участке. С этого времени начинается регулярное токование, которое приобретает всё более интенсивный характер. Самки в это время держатся поблизости, но регулярное посещение ими токовищ начинается обычно с появлением первых проталин.
Главный период токования, во время которого происходит спаривание, короток. Его длительность в нормальных условиях составляет 7–10 дней (Теплов, 1947; Pirkola, Koivisto, 1970). Этот период имеет наиболее твёрдые сроки и менее всего подвержен влиянию погоды, хотя и здесь погодные условия могут несколько растянуть его или сжать (в пределах 2–3 дней). В иное время непогода оказывает заметное влияние на ход токования, особенно ненастье с сильным ветром. Птицы могут угадывать его наступление, реагируя резким снижением активности токования или даже его прекращением за 8–12 часов до ухудшения погоды.
В разгар токования самцы собираются на токовище с вечера, до захода солнца — приходят пешком или прилетают из ближайших окрестностей, где проводят на кормёжке день и отдыхают. Прибывающие на ток самцы, прежде чем начать вечернее пение, совершают несколько демонстрационных полётов. Начав пение, они обычно уже не меняют места и поют на одном и том же дереве до глубоких сумерек, нередко ещё успевая покормиться хвоей перед сном. На землю вечером не спускаются. Утреннее пение начинается в полной темноте (на 60° северной широты — в 3 часа утра), причём самцы почти сразу же начинают петь с большой интенсивностью, исполняя песню за песней почти без перерыва.
С первыми признаками рассвета, при освещённости в 3–5 люкс на ток прилетают самки, по 1 или по 2–3, и садятся неподвижно на деревья между токующими самцами. Одновременно с этим или раньше самцы спускаются на землю характерным демонстрационным полётом, каждый на свой участок, где продолжают токовать. Самки постепенно, одна за другой, начинают садиться на землю и идут к избранным ими самцам, причём каждый перелёт самки вызывает заметное повышение активности токующих самцов. Распределяются самки крайне неравномерно; у одного самца может собираться одновременно до 4–6 самок, у другого — ни одной. Границы токовых участков в это время строго соблюдаются. Обычно за утро самка спаривается с избранным самцом один раз, но при содержании в вольерах наблюдались повторные спаривания тех же партнёров через 20–30 минут, а в течение всего периода токования каждая самка спаривалась неоднократно (Кутовая, 1976). С восходом солнца самки начинают разлетаться, часть самцов, особенно молодые, покидает токовище вслед за ними, но наиболее активные самцы продолжают токовать ещё некоторое время на земле, а затем на деревьях. На нетронутых человеком токовищах в разгар токования его окончание может затягиваться до 9–10 часов утра.
После того, как самки перестают посещать ток, самцы собираются здесь ещё довольно долго, в течение 3–4 недель, но активность токования снижается с каждым днём. В это время они прилетают на токовище задолго до заката и подолгу кормятся на тех деревьях, где ночуют. Они уже не слетают по утрам на землю и на зорях поют всё менее активно, далеко не всегда принимая при пении токовую позу. В конце токования песня вообще может исполняться при опущенном и сложенном хвосте. Как на севере, так и на юге ареала окончание пения самцов совпадает с массовым распусканием и ростом берёзовых листьев, хотя в холодные, запоздалые весны пение самцов можно слышать до начала — середины июня.
Для глухаря известно также летнее, осеннее и даже зимнее токование. Летнее токование характерно для годовалых птиц, собирающихся на весенние токовища, и происходит в июле — начале августа. Самцы при этом поют и принимают токовые позы (Кириков, 1952; Кутовая, 1976). Осеннее токование свойственно взрослым самцам, наиболее хорошо выражено в южной части ареала и может происходить с середины августа до начала ноября. Птицы поют непериодически, в разное время и в разных местах, в том числе и на токовищах. Они могут петь и в смешанных стаях, состоящих из самцов и самок. Осенью же (сентябрь) происходит незначительное увеличение семенников, но активного сперматогенеза в них не обнаружено. Зимнее токование — более редкое явление и протекает так же, как и осеннее, хотя в вольерах молодые глухари токуют всю зиму (Кутовая, 1976). Вероятно, правы те исследователи (Wiirm, 1885; Кириков, 1952), которые считают, что самцы стремятся токовать всё время, и только интенсивная линька и зимние морозы прерывают эту активность.
Территориальное поведение самцов явно зависит от их возраста и заметно меняется в ходе токования, максимально проявляясь в возрасте 3 лет и старше непосредственно в период спаривания. В это время самец особенно активно охраняет свой участок, размеры которого в среднем занимают около 0.6 га (радиус около 40 м). Места токования (токовища), как правило, отличаются большим постоянством и могут функционировать в течение сотни лет и более, однако многие токовища существуют далеко не столь длительное время. Местом тока избирается обычно участок разреженного леса (на большей части ареала соснового или с преобладанием сосны), с минимальным количеством подлеска и подроста, с вересковым или моховым покровом. Отсутствие подлеска позволяет просматривать местность вокруг по крайней мере на 50–70 м.
Часто во время токования почва на токовище почти сплошь залита водой либо же представляет пропитанный водой торфяник. В южной части ареала токовище может находиться и в лиственном лесу (дубняки, осинники, березняки) и в ельниках (Альпы, Карпаты, Алтай). Размеры токовища также весьма различны и зависят от численности птиц в данном районе и интенсивности охоты на токах. При благоприятных условиях площадь крупных токовищ может достигать 1,5 км, и здесь собирается до сотни и более самцов (Теплов, 1947). В местах, где численность низка или имеется сильный охотничий пресс, средние размеры токовища обычно не превышают 10 га, и здесь собираются 2–7 птиц. В местах, совершенно разорённых охотой, самцы могут токовать и в одиночку — явление явно вторичное и в целом виду не свойственное.
Важно отметить, что нередко в районе обитания микропопуляции имеется несколько токовищ, причём отдельные самцы могут поочерёдно появляться то на одном, то на другом из них (Теплов, 1947).
Брачный ритуал глухаря довольно сложен и состоит из ряда элементов, основным среди которых является совершенно особая брачная песня. Она сравнительно тихая (максимальная слышимость 150–200 м) и состоит из 2 частей. Общая длительность её около 5,5 секунд. Первая часть состоит из громких двойных щелчков, которые, все убыстряясь, следуют друг за другом, пока не сольются в сплошную короткую трель, тянущуюся 0,5-0,8 секунд. Трель обрывается короткой паузой (0,1–0,2 секунд), вслед за которой следует серия звонких шипящих звуков, так называемое точение, длительностью около 3 секунд. В зависимости от степени возбуждения самца, начальная часть песни (двойные щелчки) может быть то короче, то длиннее. На конец «трели» и начало «точения» накладывается ещё один, глухой, звук, слышимый только с близкого расстояния (до 15 м) и удачно сравниваемый со стуком гороха, сыплющегося в деревянное корыто (Lumsden, 1961; Пукинский, Роо, 1966). На этот звук в свою очередь накладывается несколько тихих, отрывистых «капельных звуков», напоминающих звук падающей капли: 3–5 у южно-сибирских глухарей; 5 двойных у шведских (Hjorth, 1970); 0-3 в Ленинградской области и Карелии (Потапов, 1985).
У самцов подвидов urogallus, major, aquitannicus один из этих звуков, падающий как раз на короткую паузу между «трелью» и «шипением», резко усилен, и является самым громким звуком песни, получив у немецких охотников наименование «хауптшлаг» — «главный удар» (французское и английское его наименования более удачны — они показывают сходство этого звука с тем, который производится при откупоривании бутылки). Примечательно, что этот звук свойствен только 3 указанным подвидам, весьма близким между собой (Потапов, 1971).
Одна из самых ярких особенностей песни глухаря — потеря птицей реакции на звук (глухота) между последним моментом второй части песни (трели) и окончанием третьей (точения). В это время птица, как правило, не реагирует на посторонние звуки, что с древних времён используется человеком при охоте на токующих самцов. Причина этой временной глухоты экспериментально не выяснена. Большинство исследователей сейчас склоняется к мнению, что глухота эта не следствие закрывания слухового отверстия той или иной структурой, а результат своеобразного аффекта, в котором пребывает самец при исполнении самой азартной и напряжённой части песни. Зрительной осторожности при этом птица не теряет.
Во время пения самцы могут принимать самые различные позы, но обычно при этом шея поднимается и вытягивается вертикально, голова тоже поднимается вверх, а хвост полностью раскрывается и также поднимается вертикально. Крылья слегка распускаются и отводятся немного в стороны и вниз. Из других элементов брачного ритуала особенно характерны так называемые подлёты — своеобразные демонстрационные полёты, выполняемые обычно вечером, когда самцы занимают свои участки, возвращаясь с мест дневных кормёжек. Такой полёт совершается по прямой, обычно с дерева на дерево, в сильно замедленном темпе, с усиленными громкими взмахами крыльев, нередко задевающих при этом за ветки. Заканчивая полёт, самец садится на ветку и сразу же издаёт особый трёхсложный «хрюкающий» звук «у.. .у-у».
Расстояние, проделываемое самцом при таком полете, в среднем около 50 м; птица летит на высоте вполдерева и, как правило, совершает этот полет в пределах своего токового участка. Ещё один элемент — токовой взлёт. Эти взлёты совершаются самцом на земле, причём особенно часто в период кульминации тока. Выполняя этот взлёт, птица полого взлетает на высоту 1,5–2 м и так же полого опускается. Токовой взлёт сопровождается громким хлопаньем крыльев, причём птица делает несколько взмахов на взлёте и на спуске, пролетая при этом от 3 до 7 м, и иногда меняет направление взлёта на 45–90°. В верхней точке взлёта птица на доли секунды замирает с раскрытыми полностью крыльями. В начальный период токования самцы принимают ещё особую позу «распластывания»: птица ложится на снег с раскрытыми крыльями и хвостом и находится в такой позе неподвижно несколько секунд. В результате на снегу остаётся иногда полный отпечаток птицы.
Драки между самцами могут иметь место и в начальной фазе токования, и, особенно, в его разгар. Дерущиеся самцы наносят друг другу удары крыльями, сталкиваясь при этом грудью, а также стараются достать шею и голову противника клювом. Шум от ударов крыльями во время такой драки бывает слышен на 150–200 м. Известны случаи, когда драки заканчивались гибелью одного из соперников (Boback, 1966). После окончания токования самцы начинают линьку и все лето держатся очень скрытно, проводя время в основном на земле в густых зарослях по краям болот, вырубок или гарей.
Самки устраивают гнёзда недалеко от токовищ, причём иногда могут гнездиться в непосредственной близости одна от другой, в 40–50 м (Долбик, 1961). Гнездо устраивается на сухом месте и обычно прикрыто сверху (еловой ветвью, упавшим стволом и тому подобном), но может располагаться и открыто. Гнездо — небольшое углубление в почве, выстланное сухим растительным материалом. Как исключение, гнёзда могут устраиваться на деревьях, в старых гнёздах других птиц (Ваппегтап, 1963; Василенко, 1971). В северных частях ареала известны случаи, когда самка начинала откладку яиц прямо на снег (Pulliainen, Rajala, 1973). Среднее количество яиц в кладке 7–8, но вообще это число варьирует в пределах от 4 до 16, находясь в явной зависимости и от возраста самки, и от степени её упитанности в момент размножения. Так, в одной и той же местности, в зависимости от характера зимы и весны полные кладки могут содержать то по 12, то по 5 яиц (Кириков, 1952).
Средний размер яиц в Печоро-Илычском заповеднике 58,4 × 41,2 (56,2-60,5 × 40,5-42,1) (Теплов, 1947).
Толщина скорлупы в средней части яйца 0,4 мм. Средняя масса свежего яйца (n = 29) 53,2 (Семёнов-Тян-Шанский, 1959).
Окраска варьирует очень сильно. Наиболее типичен палево-песочный или слегка коричневый фон, по которому густо разбросаны мелкие темно-коричневые пятна округлой формы (среди них обычно бывает несколько пятен более крупного размера, диаметром до 3–4 мм). Пятна разбросаны по всей поверхности равномерно. В процессе насиживания окраска яиц может сильно выцветать. Яйца начинают откладываться самкой через 3–4 дня после спаривания, с интервалами от 30 до 48 часов (Кутовая, 1976). Насиживание начинается после снесения последнего яйца и продолжается, в зависимости от интенсивности насиживания, от 22 до 30 суток, в среднем же 25–26 суток (Hoglund, 1952; Семёнов-Тян-Шанский, 1959; Ивантер, 1965; Pulliainen, 1971b). Температура наседного пятна самки при насиживании 39,6–41,2 °С. Колебания температуры яиц составляют от 30,0 до 39,3 °С, в среднем 36,3–36,4 °С (Valanne, 1966; Семенов-Тян-Шанский, Брагин, 1969). Самка отлучается на кормёжку в среднем 2–3 раза в сутки (иногда до 4–5), продолжительность отлучки от 20 до 34 минут. Яйца поворачиваются самкой от 14 до 22 раз в сутки. Самая низкая температура яйца, не повредившая нормальному развитию зародыша — 14,6 °С, была зафиксирована после 80-минутной отлучки (Lennerstedt, 1966).
Покидая гнездо, самка или взлетает прямо с него, или отходит на 1–2 м. Возвращаясь, она приземляется в 3–5 м от гнезда и затем идёт к нему пешком, внимательно оглядываясь. Вылупление птенцов происходит дружно, в течение нескольких часов, хотя в отдельных случаях между первым проклёвом на яйце и вылуплением может пройти около 2 суток (Krott, 1966; Василенко, 1971). Масса вылупившегося птенца 33–45 (Ефремов, 1940; Крутовская, Крутовская, 1953; Hoglund, 1955; Семёнов-Тян-Шанский, 1959; Ивантер, 1965).
В первые сутки масса может даже падать, поскольку первые два дня птенцы существуют в основном за счёт желточного мешка. Птенцы, однако, с первого дня активно ищут и поедают насекомых, которых они склёвывают с нижней поверхности стеблей и листьев. Температура тела у только что вылупившегося птенца 37,9 °С. Она постепенно поднимается и достигает 41,6 °С в возрасте 18 дней, когда птенцы вполне приобретают способность к терморегуляции. В первые же дни птенцы остывают очень быстро. Чем ниже температура воздуха, чем больше влажность, тем быстрее охлаждаются птенцы, тем чаще и дольше они обогреваются самкой и тем меньше корма они успевают найти, особенно потому, что в плохую погоду уменьшается и количество активных насекомых. При затяжном холодном ненастье такая ситуация приводит к гибели птенцов от голода.
Только в возрасте 14— 18 дней птенцы способны оставаться при температуре воздуха 14 °С 30 минут без снижения температуры тела и кормовой активности (Hoglund, 1955). Таким образом, первые две недели жизни — наиболее критические в жизни птенцов глухарей, так же, как и у остальных тетеревиных птиц. Но и в более зрелом возрасте — полтора месяца и даже более — во время затяжных дождей, молодые птицы, несмотря на подкормку, вдвое снижали суточные привесы и замедляли линьку (Кирпичёв, 1968). Первый месяц выводок проводит обычно в ельниках, по окраинам вырубок и, особенно, верховых болот, где более всего предпочитается полоса тростников вдоль невысоких грив, заросших лесом. Птенцы в возрасте 8 дней уже могут взлетать на высоту до 1 м. Распадание выводков начинается с 80-го дня, с появлением агрессивности у самцов и их попыток покрывать самок. Остатки выводков, состоящие из молодых самок и старой птицы, к зиме сбиваются в стайки, самцы образуют отдельные группы.
На севере ареала первые выводки появляются в конце июня (Кольский полуостров, — Семёнов-Тян-Шанский, 1959), в середине июня (Кировская обл., Северный Урал, — Портенко, 1937, Ефремов, 1940) или даже в конце мая (Свердловская обл., — Данилов, 1975). В Ленинградской области появление первых выводков отмечается в начале — середине июня (данные автора). В целом же по ареалу сроки появления вида довольно близкие, если учесть влияние погодных условий, — первая и вторая декады июня.
Вне периода токования мало заметны. Отличаются большой осторожностью и стараются незаметно уйти пешком или улететь задолго до приближения человека. Слух и зрение отлично развиты. Несмотря на большие размеры, взлетают сравнительно легко, однако полёт мало маневренный и нередки случаи, когда птица задевает крылом за сучки, особенно перед посадкой. Поэтому взлетев, глухарь, особенно самец, старается набрать высоту и подняться над деревьями, но при этом не поднимается выше нескольких метров над вершинами.
Только в осеннее время, во время спонтанных перемещений глухари могут набирать значительную высоту (Семёнов-Тян-Шанский, 1959). Полёт ровный, с равномерными взмахами крыльев, фазы скольжения кратковременны, в основном хорошо выражены перед посадкой. По земле передвигаются быстро и легко, ловко пробираясь в самом густом буреломе. Зимой на большей части ареала проводят значительную часть времени в подснежных камерах, днем кормятся, причём самцы предпочитают кормиться, расхаживая по снегу, самки же — на деревьях. Как и большинство тетеревиных птиц, глухари в зимнее время, находясь под снегом, могут подпустить человека вплотную и внезапно вылететь из-под самых ног.
Голос самцов и самок различен; помимо голосовых реакций, свойственных периоду спаривания (см. ниже) для самцов характерен особый скрипучий звук, которым птица реагирует на вспугнувшую её собаку. Потревоженная самка издаёт отрывистые звуки «ак. . ак. . .ак». Глухари, даже потревоженные, редко подают голос, будучи крайне молчаливыми.
Благодаря своим размерам, глухарь хорошо отличается в природе от остальных тетеревиных птиц. Самка глухаря на некотором расстоянии может быть спутана с самкой тетерева, но хорошо отличается от неё отсутствием белого зеркала на крыле, что особенно заметно в полёте. В ряде районов Восточной Сибири, где встречаются оба вида глухаря, бывает трудно различать в природе самок, но на близком расстоянии самки глухаря хорошо отличаются преимущественно рыжей окраской грудного пластрона. Окраска самцов резко различна, будучи почти чёрной у каменного глухаря. Молодые же самцы в первую осень могут быть легко спутаны, и здесь главным отличием является отсутствие у обыкновенного глухаря крупных белых пятен на крыле и кроющих перьях хвоста.
При благоприятных условиях зимние стаи самцов могут насчитывать до 70, а самок даже до 100 птиц. В значительной степени это разделение обусловлено различными местами обитания — самцы предпочитают более открытые биотопы, самки же держатся в более густых насаждениях. Там где численность, невысока, зимние группы самцов насчитывают обычно 3–7 птиц, причём нередко вместе с ними держатся 1–2 самки.
Образ жизни глухарей в середине зимы на большей части ареала довольно однообразен. Ночуют они в подснежных камерах, таких же, как и у остальных тетеревиных птиц, но нередко, особенно при слабых морозах, ночуют и в поверхностных лунках, а во время оттепелей — на деревьях. Птицы покидают камеры, когда уже достаточно рассветёт и либо сразу же начинают кормиться, расхаживая по снегу и объедая хвою с низких сосенок, либо перелетают на находящиеся поблизости кормовые деревья — сосны или кедры, хвоя которых особенно привлекает птиц. В результате длительных кормёжек на таких деревьях может поедаться вся хвоя и они впоследствии гибнут. Регулярная кормёжка на отдельных болотных соснах, в процессе которой крона дерева как бы подстригается птицей, приводит к формированию характерной плоской вершины. В середине дня птицы обычно отдыхают на деревьях или на поверхности снега около 2 часов и затем вновь кормятся до начала вечерних сумерок.
При сильных морозах или ветре глухари во время дневного отдыха вновь зарываются в снег. На ночёвку птицы могут расположиться тут же, у самых мест кормёжки, или же перелетают на новое место, обычно не далее 500 — 1 000 м. В сильные морозы, особенно на севере ареала, глухари кормятся все светлое время суток, без перерыва в середине дня, причём длительность этой кормёжки, при особенно низких температурах, сокращается до 1,5 часов (Теплов, 1947; данные автора). Минимальная температура, при которой наблюдалась кормёжка глухаря, — 36,5 °С (Семёнов-Тян-Шанский, 1959). Сведения о том, что при температуре ниже —40 °С глухари могут по нескольку дней оставаться в камере, не выходя на кормёжку, документально не подтверждены. В случае образования твёрдого наста, что бывает чаще всего в марте, птицы не пытаются его пробивать и ночуют на его поверхности открыто или под нависающими ветками елей.
Ювенальная линька заканчивается к месячному возрасту. Последними у самцов вырастают тёмные перья в центре груди, на шее и голове, называемые иногда (Крутовская, Крутовская, 1953) переходным пером — это типично ювенальные перья, вырастающие самыми последними. К 35-м суткам начинают расти рулевые и кроющие перья хвоста и 4-е взрослое дефинитивное маховое. Взрослые перья на груди у самцов с характерным зелёным отливом появляются на 45–50 суток. Окончательная смена всех ювенальных перьев происходит в возрасте более 100 суток и заканчивается к концу сентября. От ювенального наряда остаются только 9-е и 10-е первостепенные маховые, а у птиц из поздних выводков иногда и самые внутренние второстепенные.
Третья линька, летняя, охватывает обычно только переднюю часть тела, в основном голову, шею и грудь. Она протекает в короткие сроки (у самцов в конце мая — июне) и целиком накладывается на четвертую, полную (вторую осеннюю) линьку. Последняя начинается у самцов с выпадания 1-го первостепенного махового (конец мая). У размножающихся самок летние перья растут только после вылупления птенцов, а у самок из северных частей ареала (Северный Урал) летних перьев вообще не удалось обнаружить.
Четвёртая (вторая осенняя) линька — полная. У самок её начало задерживается до конца насиживания. Линька эта протекает обычным для тетеревиных птиц порядком и укладывается по времени в промежуток между выпаданием 1-го и отрастанием 10-го первостепенного махового пера. Синхронизация и последовательность линьки отдельных партий перьев сильно варьируют, как у разных подвидов, так и индивидуально. Так, второстепенные маховые могут линять от 2-го к 20-му, а 1-е выпадает вместе с 7-м или 8-м, либо же они линяют от 3-го к 20-му, а 2-е и затем 1-е выпадают соответственно позднее, или же появляются добавочные центры линьки среди проксимальных второстепенных маховых.
Некоторые птицы иногда не успевают сменить последние второстепенные маховые, остающиеся на второй год. Линька протекает наиболее интенсивно в августе, причём в разгар линьки самцы на короткое время даже теряют способность к полёту (Федюшин, Долбик, 1967), во всяком случае они взлетают очень неохотно и держатся в это время в наиболее укромных и глухих местах, обычно в зарослях (нередко тростниковых) по окраинам болот.
Подробнее о сокращениях можно прочесть здесь.
Песня
…
Позывка
…
Электроманок
…
Образ жизни. Wurm, 1885 : 1—340; Мензбир, 1902 : 174—242; Teплов, 1947а : 3—76; Кириков, 1952 : 70—118; Воbаск, 1966 : 1—118.
Самый крупный представитель семейства. Самцы отличаются грузным телом, сравнительно длинной шеей и крупной головой с мощным клювом. Надклювье загнуто крючком вниз. Крылья типичные для лесной куриной птицы, сильно закругленные. Хвост длинный, рулевые перья широкие с прямо срезанными вершинами. Самка по. размерам значительно меньше самца, более пропорционально сложена, с более короткими шеей и хвостом. Относительные размеры клюва также невелики. Форма крыла примерно такая же, как у самцов. Клюв у самцов светлый, нередко желтоватой окраски, у самок — черного цвета. Пальцы темно-рогового цвета, радужина коричневая.
Взрослый самец (от двух лет и старше). Голова и шея темно-шиферного цвета, с неясной темной пятнистостью, образованной как наствольными, так и поперечными черноватыми полосками, нередко с белесоватым струйчатым рисунком. Ушные перья буроватого цвета. Перья «бороды» заостренной формы, черные с зеленоватым отливом по краям (рис. 92).

Изредка по краям «бороды» попадаются перья либо белые целиком, либо же с крупными белыми пятнами (возможный атавизм — остаток белого окаймления черного горла у предковой формы).
Верх тела отличается мелким струйчатым рисунком светло-серого цвета по черному или черно-коричневому фону. Коричневый фон особенно выражен в лопаточной области. В зависимости от густоты и ширины светло-серых поперечных полосок верх кажется то более темным, то более светлым. С возрастом эти полоски делаются уже, струйчатый рисунок — мельче, а общий тон окраски — темнее. На нижней стороне тела выделяется зеленый с металлическим отливом пластрон, охватывающий полукругом нижнюю часть шеи и грудь.
Пластрон образован широкими зелеными вершинами черно-бурых перьев (рис. 93). Перед и середина брюха У старых птиц покрыты черными перьями, с зеленовато-металлическими каймами. У более молодых птиц здесь urogallus. развит сероватый струйчатый рисунок. Задняя часть брюха до подхвостья покрыта бурыми волосовидными перьями, с широкими беловато-сероватыми вершинами (рис. 94).
Посредине брюха ряд перьев несет белые пестрины, количество которых колеблется индивидуально, хотя и наблюдается тенденция уменьшения их количества с возрастом. Белые пятна расположены на вершинах и наружных опахалах перьев, причем последние, особенно в передней части брюха, часто скрыты темными вершинами соседних перьев. Боковые партии перьев несут светло-серый или белый струйчатый рисунок по шоколадному фону, а наиболее крупные и длинные перья, прикрывающие бедра, имеют белые вершины, которые тем шире, чем длиннее перо: струйчатый рисунок большей частью развит только в предвершинной части или вообще отсутствует (рис. 95). Белые вершины охватывают обе половины, опахала неравномерно (хотя есть и совершенно симметричные по окраске перья), будучи более развиты на наружных опахалах. Иногда на концах таких белых вершин развито небольшое концевое пятно шоколадного цвета. ПМП шоколадной окраски, более светлой на узком наружном опахале: при изнашивании последние нередко становятся совсем белыми, в особенности на ПМП-5 — ПМП-8. На наружных опахалах и на верхней части внутренних нередко развита коричневая крапчатость различной интенсивности. На внешних опахалах ВМП преобладает коричневый струйчатый рисунок, распространяющийся на самых проксимальных перьях и на внутренние опахала, где у вершины развит белый крап, сливающийся иногда в сплошное беловатоеі вершинное поле. ВМП-1, а иногда и ВМП-2 часто имеют в нижней части 1—2 белых пятна, как бы остатки «зеркала». Перья крылышка однотонно коричневые. Такого же цвета перья, кроющие ПМП. Малые, средние и большие кроющие крыла, как и плечевые, имеют коричневый оттенок, создаваемый струйчатым рисунком рыжевато-коричневого цвета. Нередко в вершинной части перьев этот рисунок становится сероватым или беловатым. Нижние кроющие ПМП темно-серые, самые дистальные обычно одноцветные, на остальных есть^ белая крапчатость, а на наиболее проксимальных — крупные белые поля на внутренних опахалах. Подмышечные перья белые, на самых длинных из них — черные концевые пятна. РП черные, с белыми узорами в виде
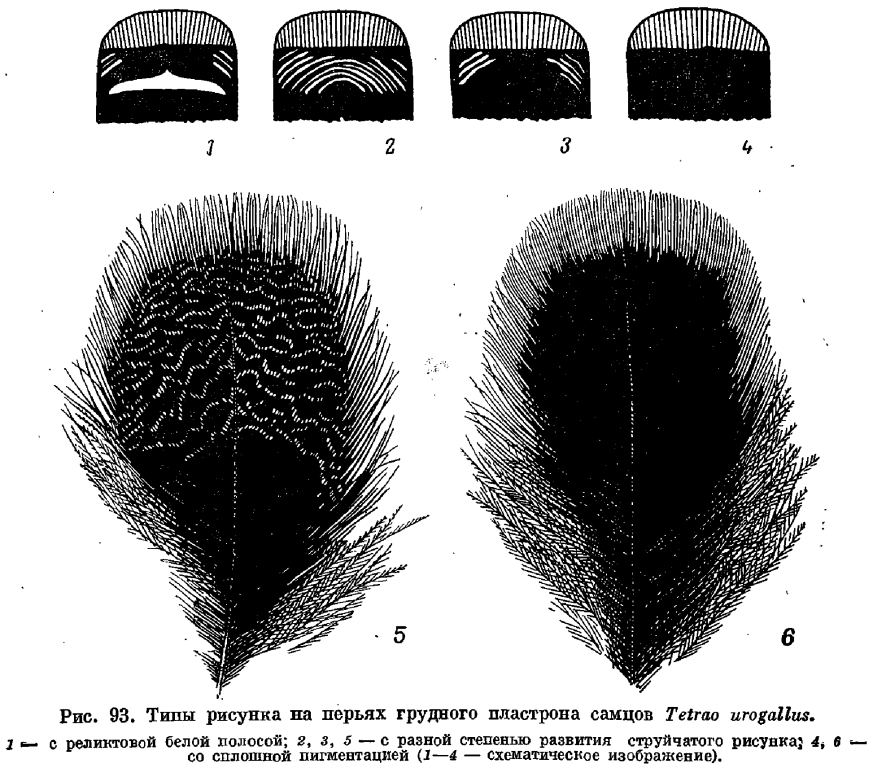
пятен и волнистых полосок в средней части пера. Узор этот лучше развит на внешних опахалах и нередко отсутствует у старых птиц на 1—3 центральных перьях. Центральная пара РП часто на 2/3 покрыта коричневым крапчатым рисунком, но вообще чем старше птица, тем этой струйчатости меньше, а у старых птиц ее может вообще не быть. ВКХ со светло-коричневым или белесым крапчатым рисунком, выраженным у старых птиц очень неясно, и вершинными белыми каймами шириной в 3—5 мм. НКХ черные, с белыми вершинами, причем на самых длинных перьях эти вершины либо испятнаны черным, либо разбиты центральной черной полосой на два вершинных пятна.
Самец в первом взрослом наряде. Отличается большим развитием белого цвета и серого струйчатого рисунка. Перья «бороды» несут в большинстве случаев неясный беловатый струйчатый рисунок, который развит также и на темно-бурых перьях, ограничивающих снизу зеленый пластрон.
На брюхе обычно много белого, гораздо больше, чем у старых птиц (за исключением южноуральского подвида), главным образом за счет развития белых предвершинных и вершинных пятен и полос, большей частью на внешних опахалах перьев. На этих же перьях нередко встречаются наствольные белые полосы и пятна. РП молодых птиц в отличие от взрослых узкие и более короткие, вершины их закруглены и несут белые концевые полосы, ширина которых заметно варьирует.

Полосы эти бывают либо на всех рулевых, либо на 2—3 центральных и одной внешней парах, причем наиболее широки они на центральной паре. Светло-серый струйчатый рисунок покрывает эту пару нередко до самого конца, а белую вершинную кайму нередко предваряет неширокая черно-бурая полоса (тип окраски РП рябчика). В средней части всех РП расположены крупные продольные белые поля сложной формы, но на крайних и центральной парах они имеют наименьшие размеры и нередко вообще отсутствуют. ВКХ также покрыты струйчатым рисунком.’ НКХ отличаются чистыми белыми вершинами. На ВМП-1, а нередко и на ВМП-2, как правило, есть 1—2 крупных белых пятна. Рыжевато-бурый крап преобладает на внешнем опахале самого длинного пера крылышка. Подмышечные перья чисто-белые. Нижние кроющие ПМП с крупными вершинными белыми полями на внутренних опахалах, с более развитым белым и рыжеватым рисунком в вершинной части пера. Плечевые перья гораздо более светлые, нередко с преобладанием струйчатого серого рисунка у вершин.
Взрослая самка. Верхняя часть тела окрашена очень пестро и представляет сочетание черных, желтовато-рыжих, белесых и белых поперечных полос. Перья здесь с широкими черными предвершинными и белесыми, с черноватым крапом вершинными полосами, часто разделенными еще узкой рыжей полоской, также несущей черную крапчатость (рис. 96). У некоторых особей на перьях спины вершинная полоса охристого цвета, с узкой более светлой каймой. В общей окраске верха спина выглядит наиболее темной, а охристые предвершинные полосы перьев особенно четко выражены на зашейке. Нижняя часть тела также пестрая. Подбородок беловатый, горло желтовато-рыжее, как- и передняя часть нижней стороны шеи. Остальная часть шеи пестрой окраски, из беловатых вершинных и черных и рыжих предвершинных полос. Грудь желтовато-рыжая, в нижней части ее имеются черные поперечные полоски, которые у некоторых экземпляров занимают весь грудной пластрон. Рыжие перья грудного пластрона несут белесоватые вершинные полоски (рис. 90). В пестрой окраске брюха доминируют широкие белые вершины перьев, временами придающие ему сплошь белую окраску. Так же окрашены и бока тела, но здесь вершины перьев часто испятнаны черным и не столь широки.
Внутренние опахала ПМП светло-шоколадной окраски, со светлыми каемками в верхней части пера. Внешние опахала более светлые, с нечеткими изломанными поперечными полосками светло-охристого цвета. ВМП с белыми вершинными каймами, особенно широкими на самых внутренних перьях и нечеткими охристыми полосами по светло-шоколадному фону. На внутренних опахалах этот рисунок меньше развит. Перья крылышка шоколадного цвета, с рыжеватым поперечным рисунком на наружных опахалах. Большие ВКК также шоколадной окраски, с рыжеватыми каемками. Все ВКК с белыми вершинами, особенно хорошо выраженными в проксимальной части крыла , и охристыми поперечными полосками по темно-шоколадному фону. Тип окраски нижних кроющих первостепенных маховых такой же, только более бледный — белая вершина и поперечные охристые полоски по светло-шоколадному фону.
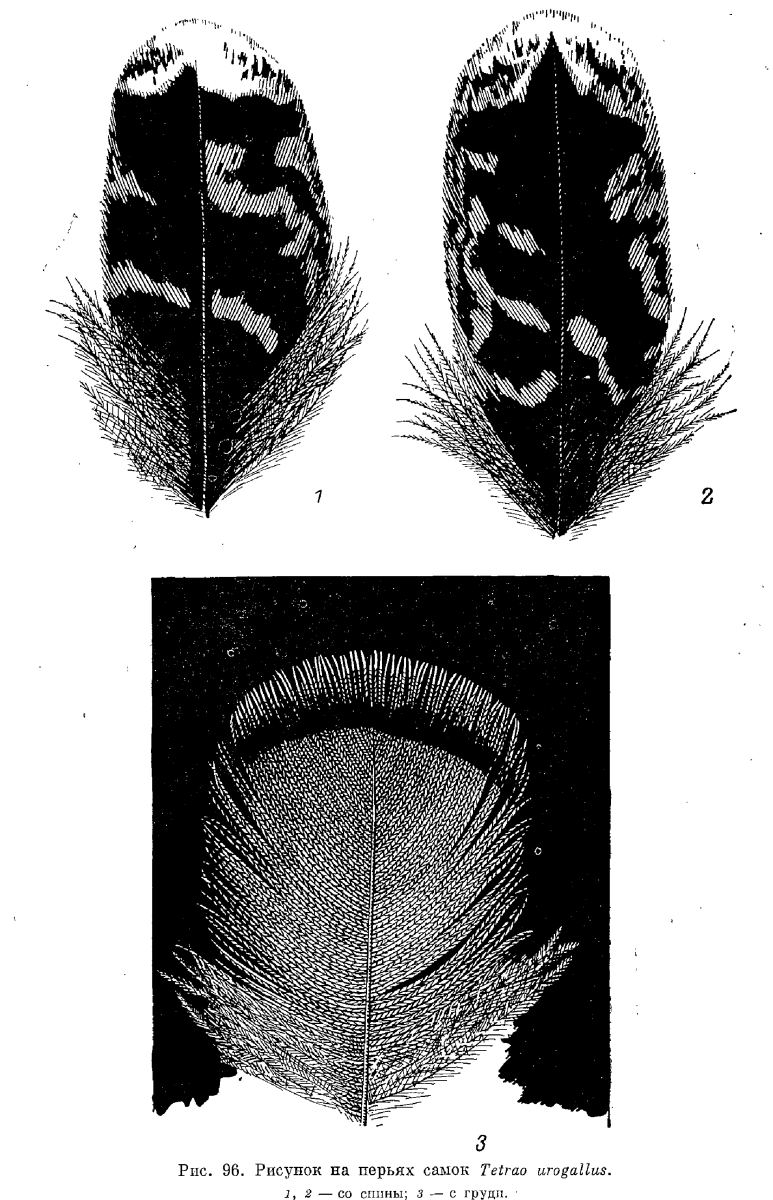
РП с белыми концевыми каймами, разрисованы широкими поперечными полосами охристо-рыжего и черного цветов. ВКХ окрашены так же. Нижние кроющие при том же типе окраски несколько бледнее, с более широкими охристыми полосами.
Самки обыкновенного глухаря имеют два основных типа окраски с полными переходами между ними — «рыжую» и «серую» фазы. В первом случае преобладает желтовато-охристая окраска, грудной пластрон чистого ржаво-охристого цвета, без темных поперечных полос. Белые каймы и вершины перьев по всему телу выражены нерезко и иногда имеют желтоватый оттенок, темные полосы обычно бурого цвета. Поперечные полосы на нижней стороне тела крупные, оперение ног светлое, с желтоватым оттенком.
Самки «серой» фазы имеют более темную окраску, бока головы темные, с резким белым и черным крапом. Грудной пластрон также испещрен четкими поперечными полосами черного и белого цветов, которые становятся здесь преобладающими. Перья плюсны темные, с мелким бурым рисунком.
Эти типы окраски встречаются у самок любой популяции, но в холодных континентальных районах преобладает «серая» фаза, а в более теплых и влажных — «рыжая». В некоторых местах, как, например, на юге Красноярского края, все молодые самки имеют первый взрослый наряд «рыжей» фазы (Крутовская, 1958), что говорит о большей филогенетической древности этой фазы по сравнению с «серой». Существенно также, что, по данным Крутовской, характер окраски взрослых самок при содержании в неволе не изменяется при последующих линьках, т.е. он генетически закреплен.
Самец в ювенальном наряде. Верх головы темно-серый, с рыжеватым оттенком над бровями. Перья здесь несут вершинную черно-бурую и светлую предвершинную полоски, нередко есть белая стержневая полоска. Над «бровями» перья имеют коричневые кончики с тонкими черными поперечными полосками. Бока головы буроватые или серовато-рыжие, с белым каплевидным рисунком, который, охватывая горло, распространяется на переднюю часть шеи. Кроющие уха серые. Горло грязно-белое, по краям его перья нередко несут узкие белые каймы, создающие чешуйчатый рисунок. Зашеек и бока шеи — в мелких поперечных полосках бурого и белесого (часто с охристым оттенком) цветов, образуемых бурыми вершинными и белесыми предвершинными полосками. Спина пестрой буровато-охристой окраски, на перьях преобладает поперечный (иногда струйчатый) рисунок из темно-бурых полос различной ширины на белесовато-охристом фоне. Особенно широки темно-бурые предвершинные полосы. Нижняя часть спины, поясница и надхвостье более серые, с белыми пятнышками: перья здесь унесут наствольную белую полосу, расширяющуюся в предвершинное и вершинное пятна. Грудь рыжевато-бурая с более темной буроватой серединой. Наблюдаются два типа окраски груди (кол. ЗИН АН СССР, 9 экз.). Один тип свойствен подвиду Т.u. uralensis (сюда же относится один экземпляр Т.u. taczanowskii) и отличается наличием в центре грудного пластрона серовато-бурых перьев с белым струйчатым рисунком и (на части перьев) с широкими вершинными или предвершинными полосами. Другому типу окраски (подвиды Т.u. pleskei, Т.u. obsoletus, Т.u. taczanowskii) свойственно очень темное бурое пятно в центре грудного пластрона, слегка испещренное узкими белыми наствольными полосками. Брюхо грязно-серого цвета, с неясными буроватыми пятнами. Перья на боках светло- или темно-бурые, с рыжеватой поперечной пойосатостью, носящей часто струйчатый характер, с концевым белым пятном. ПМП темно-бурые, с волнистым коричневым рисунком на наружном опахале и верхней части внутреннего.
Сходно окрашены и ВМП, но поперечный рисунок на внешнем опахале более четкий, образован желтыми полосками. Нередко внешнее опахало окаймляет желтоватая полоса. На проксимальных ВМП рисунок усложняется за счет краевых волнистых темно-бурых пятен и распространяется также на внутреннее опахало. Для этих же перьев характерна либо беловатая вершина, либо предвершинное наствольное каплевидное пятно такого же цвета, либо же вершинная наствольная полоска, расширяющаяся на конце в вершинное пятно.
Большие, средние и малые кроющие крыла охристо-коричневые, с неясным более светлым поперечным рисунком, легкой бурой струйчатостью и беловатой яаствольной полоской, кончающейся вершинным наствольньгм пятном. На проксимальных перьях рисунок усложняется бурыми пятнами и полосами, как ш на внутренних ВМП. РП в верхней части поперечно-полосатые, с белыми конечными каймами и чередующимися светло-желтоватыми и буровато-коричневыми полосами. В нижней части перьев рисунок менее четок и правилен.
Самка в ювенальном наряде отличается большим развитием рыжих тонов в оперении, в особенности на зашейке, шее, боках головы и ушных перьях, а также чистым желтовато-рыжим цветом на груди.

РП сохраняют правильный поперечно-полосатый рисунок, доходящий почти до самого основания. Несколько иначе окрашены нижние кроющие перья первостепенных маховых: на них гораздо большее развитие получает, начиная с самого дистального пера, поперечно-полосатый рисунок из беловатых и желтоватых полос и пятен, а на самых проксимальных перьях развиты белые поля, тянущиеся вдоль стержня и внутреннего края опахал. Различий в окраске маховых перьев самцов и самок, которые находил Хеглунд (Hoglund, 1952), на нашем материале обнаружить не удалось.
Пуховой наряд. По Хеглунду, пуховые птенцы могут быть различимы по полу благодаря различной окраске клюва, но все 8 пуховых птенцов в коллекции ЗИН АН СССР имели окраску клюва совершенно одинаковую. Отрицают возможность различать пол у пуховичков и авторы, занимавшиеся разведением глухарей в неволе (Крутовская, Крутовская, 1953; Krott, 1966).
Низ птенца охристо-желтоватый, с неясной охристой перевязью на груди и охристым подпалом на подхвостье. На верхней стороне тела — довольно сложный узор из черноватых и охристых полос. Почти всегда можно различить охристую полосу, с более или менее выраженным черным бордюром поперек верхней части спины, охристую полоску на пухе, кроющем крыло и более темноватую, с неясным бурым окаймлением полосу, идущую от межлопаточной области к копчику. Узор на голове (рис. 97) представлен черными пятнами у основания клюва,- над «бровями», у глаза и около ушного отверстия, неясными черно-бурыми штрихами на голове с неясной окантовкой сзади, несколькими полосками на затылке и V-образной полосой на лбу, направленной вершиной назад. Последняя у некоторых экземпляров может быть представлена в виде трех отдельных пятен. В целом можно наметить контуры «шапочки» на голове, с сохранившимся сзади окаймлением и черной срединной полосой, идущей от «шапочки» назад. Плюсна опушена до самых пальцев, и пух заходит на бока среднего пальца.
Размеры и масса взрослых самцов в пределах ареала существенно варьируют, что обусловлено как географической изменчивостью, так и возрастными отличиями. Работами С.В. Кирикова, О.И. Семенова-Тян-Шанского и С.П. Кирпичева установлено, что рост глухарей происходит по крайней мере до трех лет и каждая возрастная группа отличается определенными размерами, увеличивающимися с возрастом. Картину усложняют индивидуальные отклонения, благодаря которым размеры и масса отдельных возрастных групп перекрываются. На коллекционном материале отделить двухгодовалых самцов от более старших в большинстве случаев невозможно, и данные по размерам самцов в коллекциях приводятся но двум возрастным группам — молодые птицы в возрасте до 15 мес и взрослые — старше этого возраста. Молодые самки уже в первую осень становятся трудно отличимыми от взрослых.
Самцы:
длина тела 955—992 (Кирпичев, 1960),
крыло 377—430,
хвост 247—351,
плюсна 62—87,
средний палец (без когтя) 59—77,
длина клюва 30—41.4,
ширина клюва 20.4—27.5.
Самки:
длина тела 620-675,
крыло 280—322,
хвост 163—205,
плюсна 46—59,
средний палец 46—58,
длина клюва 19—26,
ширина клюва 14.2—18.2.
Масса взрослых птиц колеблется в следующих пределах: самцы 3500—6500 г, самки 1440—2210 г. Следует учесть, что эти данные получены на тысячах птиц из самых разных популяций. Средняя же масса птиц в отдельных популяциях гораздо более стабильна и большей частью колеблется в пределах 3900—4300 г у самцов и 1700—2000 г у самок.
Масса годовалых птиц заметно меньше, особенно у самцов, и составляет в первую зиму 80 %, а у самок 91 % от массы взрослых птиц. Самцы продолжают увеличивать свою массу в процессе онтогенеза по крайней мере до 3-летнего возраста. Так, по данным Кирпичева (1961), годовалые самцы в Забайкалье имеют массу в среднем 2850 г (2620—4100), самцы в возрасте двух лет 4350 г (4120—4800), трех лет и старше 5530 г (5000—6450).
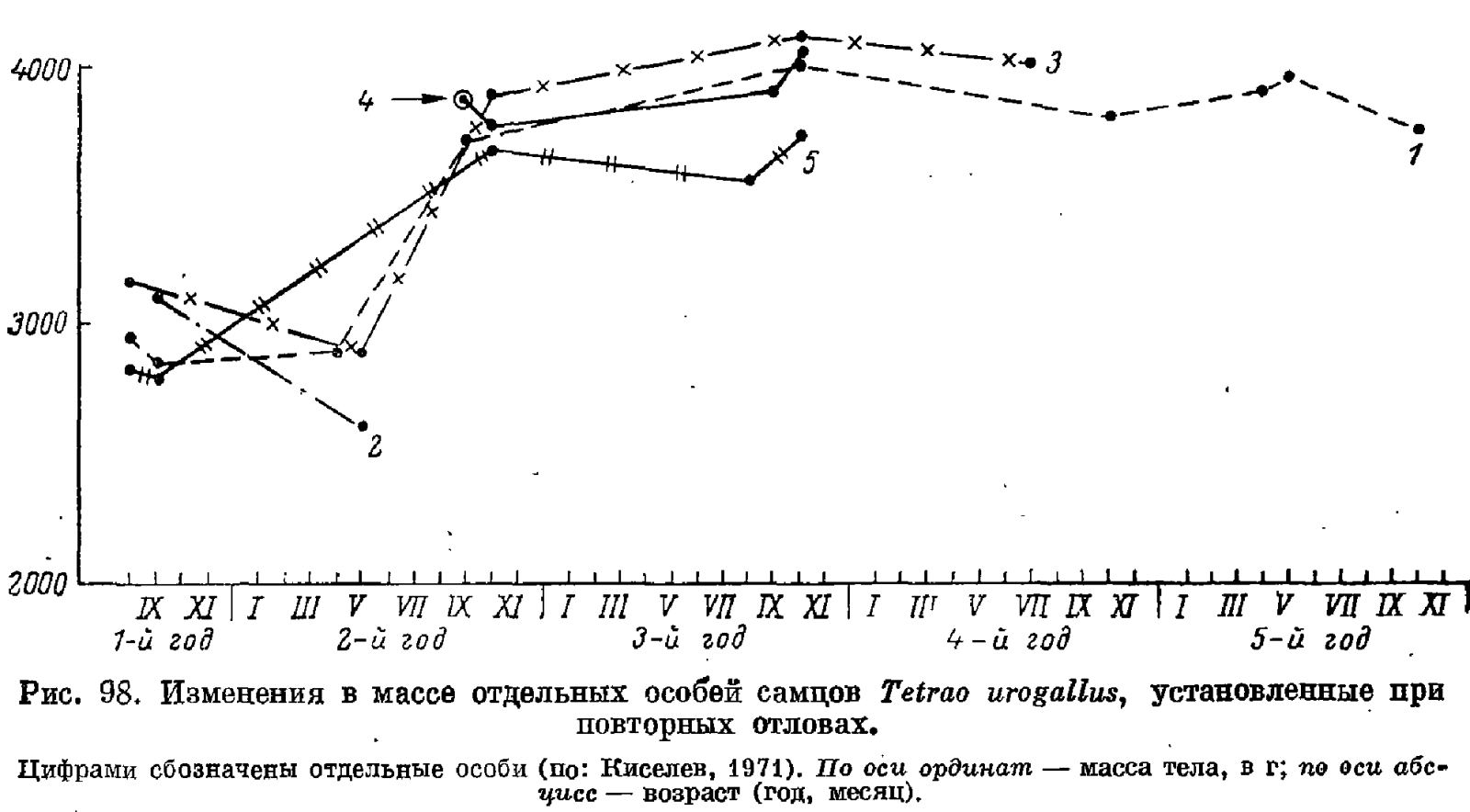
Следует особо заметить, что средняя масса птиц одной возрастной категории в одной и той же популяции может заметно меняться в разные годы в зависимости от условий среды. Уникальные данные по динамике массы глухарей в природе собраны Киселевым (1971) методами повторных отловов (рис. 98). Эти данные показывают, что начиная с поздней осени и до следующего лета рост молодых глухарей прекращается, в летние и осенние месяцы возобновляется вновь, затем опять идет период стабилизации и, наконец, в третью осень своей жизни самцы достигают максимальной массы, которая впоследствии может даже снижаться. Обращают внимание широкие индивидуальные вариации изменения массы, которые зависят и от особенностей данной особи, и от климатических условий зимовки, и от многих других факторов, в частности от зараженности гельминтами.
Максимальная масса, которой могут достигать самцы глухаря, 6500 г. Все остальные сведения о глухарях массой 7—8 кг и даже 10 кг, довольно многочисленные в охотничьей литературе, документально не были подтверждены.
Сезонная динамика массы до сих пор подробно не изучена на достаточном материале, но, видимо, она не отличается существенно от общей для тетеревиных птиц картины. Во всяком случае немногие точные данные эту картину подтверждают. Самцы достигают максимальной массы в октябре—ноябре, затем снижают ее в течение зимы и вновь немного увеличивают к началу токования, минимальная масса — в июле, во время наиболее интенсивной линьки. У самок в Мещерской низменности всегда наблюдается четкое увеличение массы весной, причем степень этого увеличения находится в прямой зависимости от суммы положительных температур данной весны. Превышение весенней массы над осенней достигает в зависимости от этого фактора от 2.44 до 9.47 % (Киселев, 1971).
Линька. Развитие оперения у глухарей в онтогенезе и линька изучены достаточно полно (Снигиревский, 1950; Крутовская, Крутовская, 1953; Семенов-Тян-Шанский, 1959; Кирпичев, 1972).
На 9-е сутки развития эмбриона появляются 4 ряда бугорков перьевых фолликулов эмбрионального пуха в пояснично-крестцовой части и несколько сосочков на бедре, 2 ряда бугорков на хвосте и крыльях. На 12-й день ряды перьевых фолликулов намечаются вдоль груди, а на средней спинной линии выражены уже очень хорошо. Конусообразные бугорки есть также на бедре, плечевой птерилии, по постаксиальному краю кисти и субаксиальной части предплечья. Хорошо заметен и яйцевой зуб. На 15-й день увеличивается число перьевых сосочков, конических и вытянутых волосовидных образований по телу. На 17-й день голые участки кожи остаются по бокам тела, на передней части шей, плюсне и висках, все остальное покрывается формирующимся пухом.
На 20-й день появляются трубки ювенальных перьев первой генерации, на верхушках которых сохраняются конусы эмбрионального пуха — это ПМП-1 — ПМП-3. Одновременно появляются и их кроющие. Между 22-м и 25-м днями появляются верхушки пеньков ПМП-4 и еще 3 кроющих, пеньки ВМП с 3 по 11 и 13 кроющих. Вылупляющийся птенец уже имеет пеньки ПМП длиной по 2—2.5 мм, всего их при вылуплении — 5.
В первые сутки у птенцов, все время обогреваемых самкой, разворачиваются ПМП-1 — ПМП-5 и ВПМ-3 — ВПМ-11, 3 пары пеньков цлечевой птерилии и внутренняя пара перьев крылышка. Появляются пеньки ПМП-6 — ПМП-7.
На вторые сутки появляются пеньки РП и перьев надхвостья. С началом самостоятельного питания отпадает птенцовый зуб и чехлик надклювья.
К 10-му дню ювенальный наряд, охватывая плечевые птерилии, развивается на спине и груди. К 11-му дню заканчивается рост ювенального ПМП-1.
К 15-му дню ПМП-1 и ПМП-2 вытесняются пеньками соответствующих маховых первоосеннего наряда. Примерно в это же время появляются пеньки ПМП-9 и ПМП-Ю и ВМП-1 и ВПМ-2, отрастают ушные перья, перья подхвостья, 9 пар РП.
К 20-му дню появляется дефинитивное ПМП-3. Ювенальное оперение покрывает все тело, за исключением головы, середины груди и брюха, где сохраняется птенцовый пух. К месячному возрасту птенцы полностью заканчивают линьку в ювенальный наряд. Последними у самцов вырастают ювенальные перья особой, окраски в центре груди («бурое пятно», см. описание ювенального наряда), а также на шее и голове (пепельно-бурые с конечным каплевидным пятном), которые Е.А. Крутовская и Е.В. Крутовская назвали «переходным» пером.
К 35-му дню начинают расти рулевые и кроющие перья хвоста взрослого наряда.
К 45-му Дню у самцов по бокам зоба появляются первые перья первоосеннего наряда с зеленым металлическим отливом (у глухарей в вольерах эти перья появлялись у самцов на 50-й день), сменяются ВМП-3, идет линька верхних кроющих крыла, плечевых, перьев спины и надхвостья. На голове перья сменяются начиная с «бороды» и темени, заканчивается смена оперения на надбровных участках, щеках и в углах рта.
На 50-й день у самцов по бокам груди видны два зеленых пятна первоосеннего наряда, шея и голова еще в ювенальном пере, на горле самцов уже видна черная «борода». К 70-дневному возрасту у самцов ювенальное оперение сохраняется только на середине шеи, у основания нижней челюсти, на щеках, над глазами, кое-где в центре зоба.
Почти все средние кроющие крыла и большая часть малых тоже ювенальные, как и ПМП-8 и 5 проксимальных ВМП. Окончательная смена всех этих перьев происходит в возрасте более 100 дней, заканчиваясь к концу сентября.
Третья по счету линька — летняя, частичная, происходит на следующий год в начале лета. Она охватывает главным образом голову, шею, зоб, верхнюю часть груди и переднюю часть спины, а также некоторые контурные перья на остальных птерилиях. Ряд перьев выпадает без последующей замены, что существенно разреживает густое зимнее оперение. Эта линька короткая. У самцов она захватывает окончание токов, проходя в основном сразу же после них, С середины или конца мая и накладывается на четвертую полную (или вторую осеннюю) линьку, начинающуюся почти одновременно с выпадением ПМП-1.
С конца июля перья летнего наряда постепенно заменяются осенними. У выводящих самок летний наряд развивается только после вылупления птенцов.
Неразмножающиеся самки линяют в сроки, сходные с молодыми самцами.
По данным С.П. Кирпичева, у размножающихся самок летний наряд развивается незначительно, в основном над «бровями» и на шее. У самок из северных частей ареала (Северный Урал) нам вообще не удалось обнаружить перьев летнего наряда. Возможно, что в условиях короткого лета с частыми понижениями темнературы летний наряд вообще не развивается.
Вторая осенняя линька, четвертая по счету, полная. Она начинается с выпадением ПМП-1, которое у самцов происходит в конце мая, а у самок задерживается до окончания насиживания, т.е. до начала—середины июня. Линька эта протекает обычным порядком и укладывается по времени в промежуток между выпадением ПМП-1 и отрастанием ПМП-10. Синхронизация отдельных этапов линьки между собой заметно варьирует. ВМП, к примеру, могут линять от 2-го к 20-му, а ВМП-1 выпадает вместе с ВМІІ-7 или ВМП-8, или же они линяют от 3-го к 20-му, а 1-е и 2-е выпадают соответственно позднее, или же здесь появляются добавочные центры, откуда линька идет в обе стороны. Можно заметить, что с возрастом птицы асинхронность линьки ВМП увеличивается.
Некоторые поздно размножавшиеся самки и старые самцы иногда не успевают сменить отдельные внутренние ВМП, и они остаются на второй год.
Разгар линьки падает на август. Есть сообщения о том, что в это время самцы даже теряют способность к полету (Федюшин, Долбик, 1967). Сведения эти пока не подтверждены, но известно, что в разгар линьки глухари действительно летают очень неохотно, держатся в наиболее укромных местах и от опасности стараются уходить пешком.
Распространение. Обыкновенный глухарь населяет зону палеарктической тайги от самых западных окраин и на восток примерно до 120° в.д. (рис. 99). Детально очертить его ареал в настоящее время очень трудно, поскольку под влиянием человека он быстро сокращается.
Северная граница распространения обыкновенного глухаря совпадает с северной границей лесов. В Скандинавии она проходит по 70° с.ш., на Кольском п-ове — по крайней мере до 69°30 (добывался весной и летом 1969 г. в окрестностях г. Никель и ст. Луотсари; С.Л. Потапов, устное сообщ.). От Кольского залива северная граница опускается заметно южнее. Глухарь населяет леса центральной части Кольского п-ова (бассейны озер Имандра, Нотозера и р. Туломы, система р. Колы, р. Умбы, окрестности Ловозера). По р. Поною глухарь встречается от истоков до пос. Каневки (67°09′ с.ш., 39°40′ в.д.).
Н а юго-востоке полуострова он есть у села Пялицы, но до Полярного круга не идет (Семенов-Тян-Шанский, 1959). К востоку от Белого моря граница начинается примерно на. широте 65° (Снигиревский, 1946), но вновь поднимается к Полярному кругу на Печоре и даже несколько заходит за него по долине p. Шапкина, правому притоку Печоры, откуда в коллекции ЗИН АН СССР есть экземпляры старых и молодых птиц.

По западному склону Урала распространен к северу, по крайней мере до 66° с.ш. (мои данные), найден на Усе (Кириков, 1975), по восточному склону он встречается до ст. Красный Камень, но гнездования здесь не наблюдалось, а ближайшие выводки были обнаружены в 80—90 км южнее (Данилов, 1975). Как далеко идет глухарь к северу по р. Оби — неизвестно, но под Салехардом по крайней мере раньше встречался (экз. кол. ЗИН АН СССР). На севере Красноярского края встречается в южных районах лесотундры, где регулярно гнездится до 67°30 с.ш., а по долинам Енисея — до 71°30 с.ш. (Яковлев, 1930). Однако, по последним сведениям (Назаров, 1968; Сыроечковский, Рогачева, 1968), глухарь по р. Таз достигает только Полярного круга, а по Енисею идет вниз лишь до устья р. Хантайки (68° с.ш.).
В долине Нижней Курейки не гнездится (Шиллингер, 1927). Далее к востоку граница точно не выяснена, видимо, она значительно спускается к югу, к долине р. Нижней Тунгузки. Во всяком случае в горах Путорана, в районе Норильских озер и р. Рыбной Кречмар (1966) встретил только каменного глухаря.
На Нижней Тунгузке обыкновенный глухарь встречается повсеместно, вплоть до истоков, но в среднем течении реки преобладает каменный глухарь. Далее к востоку обыкновенный глухарь водится по Вилюю, в особенности в верховьях этой реки и по ее притокам. Граница ареала в бассейне Вилюя точно прослежена Андреевым (1953, 1974). По бассейну Чары глухарь доходит только до нижнего правого ее притока р. Джелинда, затем граница пересекает низовья рек Большой и Малый Болтуобуй, Аппая, Вилючан, Кемпендяй — по бассейну Кемпендяя доходит до 121° в.д. и идет к югу, по бассейну притока-р. Лены — р. Наманна — к Лене. У г. Вилюйска не встречается, не переходит на левый берег Вилюя, вообще восточнее 121° в.д. его нет, несмотря на то что к востоку, в междуречье рек Лены и Вилюя, распространены те же типы леса, с участками высокоствольных сосняков. Экземпляр в коллекции ЗИН АН СССР, на этикетке которого значится «Вилюйск», видимо, был привезен сюда для продажи с юга или запада. Далее обыкновенный глухарь найден к востоку от Олекмы, в долинах рек Чара и Токко, и на северо-восточном побережье Байкала, где он есть всюду в прибрежной части. Он населяет лесной пояс Хамар-Дабана, и наиболее восточная точка в Забайкалье — устье р . Мун (Поляков, 1873), правда, не подтверждается последующими исследованиями. От южного Прибайкалья граница идет к Кяхте, где в окрестных борах глухари встречались по крайней мере до 1940-х годов (А. Н. Леонтьев, устное сообщ.), и далее к северному берегу оз. Хубсугул, откуда по южной окраине Саян идет далее на запад по хр. Танну-Ола, где глухарь населяет только северные склоны, ибо южные безлесны (Янушевич, 1952). Глухарь есть всюду в горной тайге Алтая, но в Монгольском Алтае, в лесах его северо-западной части мною не найден.
Изолированный островок ареала есть в Калбинском Алтае, в борах Шибунды и Каиндинском (Кузьмина, 1962). От западного Алтая граница идет к северу по левому берегу Оби, где глухарь еще недавно населял сосновые боры и откуда в коллекции ЗИН АН СССР есть экземпляры (из-под Барнаула), вплоть до 56° с.ш. Отсюда граница следует к западу, огибая с севера Барабинскую степь, и, видимо, спускается к югу небольшими выступами по долинам Иртыша, Ишима и Тобола. По правобережью Иртыша, в частности, есть указания о нахождении его в борах окрестностей Семипалатинска (Белокаменск) и Павлодара (Хахлов, Селевин, 1928; Селевин, 1929; Залесский, Залесский, 1931). Еще недавно глухарь встречался в борах Курганской и Челябинской областей (Санарский бор у Троицка), населял кустанайские (Полферов, 1896) и кокчетавские боры: в последних выбит в начале века (Штегман, 1934а). На Южном Урале идет далее всего к югу до 51°30′ (массив Шайтан-Тау; Кириков, 1952).
Отсюда на запад граница идет через Бузулукские боры к Жигулям, по правому берегу Волги спускается до 52°30′ с.ш. (Черкасский лесхоз; Болгов, 1968), затем, видимо, вновь поднимается севернее, к лесам Мордовской АССР (для Пензенской обл. сведений нет). В Рязанской обл. глухари обитают только в северной части (Хомяков, 1901), в Калужской обл. они были известны для Козельского, Жиздринского, Мосальского, Перемышльского районов (Филатов, 1915). Следуя отсюда к юго-западу, граница охватывает с юга леса по левобережью Десны в Карачевском и Трубчевском районах Брянской обл. (Граве, 1933; Суслова, 1937) и самые северные районы Украинской ССР, в последней этот вид находится на грани исчезновения. На Западной Украине глухари обитают в горных лесах Карпат.
В Европе, вне пределов СССР, глухарь сохранился в Польше (в воеводствах Белостокском, Ольштинском, Козалинском, Щецинском, Будгощском, Катовицком, Краковском, Вроцлавском, Жешувском, Гданьском и Люблинском, в последнем, правда, почти исчез; Marchlewski, 1962), кое-где в восточных частях ГДР, а также в горных лесах Кантабрии, Пиренеев, Альп, Карпат, Родоп, в горах Югославии и цетральной Европы (Арденны, Рейнские Сланцевые горы, Гарц, Тюрингенский Лес, Рудные горы, Судеты, Шумава, Фраконская и Швабская Юра, Шварцвальд, Вогезы). В центральной Франции, в Овернских горах истреблен в XVIII в. (Andrew, 1958). Обитал всюду на Британских островах, но был полностью истреблен в Англии в XVII, а в Ирландии и в Шотландии в XVIII в. Глухарь вновь был интродуцирован в Шотландию из Швеции в 1837 г. и сейчас к настоящему времени широко расселился по стране между Дернек Фирс на севере и долиной Ферт-Клайд на юге (Bannerman, 1963).
В СССР интродуцирован недавно в сосняках Кокчетавской обл. (см. далее).
Места обитания. Глухари обитают в самых разнообразных типах леса, различной густоты, увлажненности, возраста насаждений в зависимости от сезона и географического положения местности, но в целом по ареалу прослеживается отчетливая связь с сосной, особенно в зимнее время. Также очевидно предпочтение, оказываемое птицами лесам с разнообразной экологической обстановкой —различными по возрасту участками леса, полянами, гарями, болотами, т. е. лесам, в общем более разреженным. Сплошной темнохвойной тайги глухарь избегает. Сезонная смена местообитаний довольно отчетлива и в большинстве случаев заключается в перемене зимних местообитаний (сосновые леса) на летние (ельники).
В отношении рельефа глухарь мало привередлив. Он населяет обширные низменности, нередко сильно заболоченные, но при наличии на них возвышенностей типа сельг предпочтение отдается им. Столь же охотно населяет нагорную тайгу, где поднимается до верхней границы хвойного леса, на Алтае, в частности, до 2000 м над ур.м. (Сушкин, 1938).
Численность. Большое количество сведений о численности обыкновенного глухаря и их сравнительное однообразие (табл. 41) показывают, что наиболее обычна осенняя численность в 10—16 птиц на 1000 га. Из общего ряда цифр резко выпадают данные О.И. Семенова-Тян-Шанского и В.П. Теплова (Печоро-Илычский и Лапландский заповедники), А.Н. Романова (бассейн Кожвы), И.В. Измайлова и Е.И. Павлова (междуречье Нижней Тунгузки и Чоны) и некоторых других, превышающие остальные в 10—20 раз, но хорошо согласующиеся между собой. Эти данные показывают ту максимальную плотность, которой может достичь вид в особо благоприятных условиях — при максимуме численности и отсутствии отрицательного влияния антропогенных факторов. Возможно также, что при осенних концентрациях глухарей на ягодниках в годы высокой численности плотность птиц может быть еще выше.
Колебания численности глухарей изучены недостаточно. Можно только отметить, что в южной половине ареала численность более стабильна, чем в северной. Это находит свое объяснение црежде всего в характере климатических условий, непосредственно влияющих на успех размножения. Исследования Хеглунда (Hoglund, 1955) в Швеции показали исключительную важность погодных условий в первую декаду жизни птенцов. Позднее эти данные были подтверждены Даниловым. (1975) на Урале, где наибольшие корреляции численности глухарей со среднедекадными температурами наблюдаются для второй декады июня (-0.534 +0.036), т.е. как раз во время вылупления и первых дней жизни птенцов, Поэтому вполне понятна большая амплитуда колебаний численности в северной части ареала: погодные условия в июне там менее стабильны и длительные похолодания с затяжными дождями способны погубить почти весь приплод данного года.
В случае же благоприятной погоды численность будет возрастать заметно быстрее, чем у южных популяций, благодаря меньшей смертности в выводках у северных птиц. Так, размах колебаний численности глухарей в средней части Урала достигает б-кратного размера (Данилов, 1975).
О преимущественном влиянии на численность глухарей именно погоды хорошо говорит неодновременность пиков численности даже в соседних областях. Например, за 10 лет наблюдений (с 1948 по 1957 гг.) численность глухарей на Северном Урале была высокой в 1948, 1949, 1950, 1953, 1955 и 1956 гг., тогда как на Среднем Урале высокая численность наблюдалась только в 1950 и 1956 гг., а в 1948, 1953 и 1955 гг. она была низкой (Данилов, 1975).
В условиях средних широт восточноевропейской лесной зоны (50—60° с.ш. — Ленинградская, Вологодская, Калининская, Новгородская и другие области, а также Эстонская ССР) численность глухарей отличается гораздо большей стабильностью по сравнению с таковой летальных тетеревиных.
В последние годы на многие популяции глухарей резко возросло отрицательное влияние антропогенных факторов, среди которых особенно выделяются три — загрязнение среды, факторы беспокойства и охота. Влияние различных хищников на численность глухарей явно не столь значительно, как у остальных видов семейства, благодаря крупным размерам птиц. Взрослые самцы в сравнительно редких случаях делаются добычей куницы, соболя, лисицы, волка, рыси, а из птиц — ястреба-тетеревятника, беркута, орлана-белохвоста, сапсана, кречета, филина. Из них наиболее часто добывают взрослых птиц куница, беркут и филин. Список хищников, наносящих урон поголовью птенцов и гнездам, значительно более обширен, он включает ряд мелких куньих, канюков (обыкновенного и зимняка), ястребов и соколов, а на первом месте оказывается лисица. В последнее время в ряде областей западной части ареала, особенно в Западной и Центральной Европе большой урон численности глухарей наносит кабан, ставший здесь очень многочисленным и разоряющий большое число гнезд.
Популяционный состав. По данным Теплова (1947а), в Печоро-Илычском заповеднике соотношение полов в популяциях близко к 1 : 1 — самцы составляют 58 %, самки — 42 %. Соотношение же полов в выводках несколько иное — 45 % самцов и 55 % самок. Ряд данных свидетельствует б том, что среди птенцов самки преобладают, но из-за более высокой их смертности в старших возрастных категориях становится больше самцов. По другим данным, в птенцовом возрасте гибнет больше самцов (Linden, 1983b). В Карелии среди молодых птиц самки составляют 57.3 %, среди взрослых — 44.8 % (Ивантер, 1974). В Шотландии соотношение полов равное (Johnstone, Zwickel, 1966). Доля молодых птиц в популяциях глухарей подвержена очень сильным колебаниям в северной части ареала, изменяясь в различные годы от 3 до 81 % тогда как в подзоне южной тайги возрастной состав популяций более стабилен — молодые птицы составляют 53—80 % (Семенов-Тян-Шанский, 1959; Романов, 1975). Однако, кажется, здесь нет каких-то постоянных взаимоотношений и популяционный состав каждой конкретной популяции весьма изменчив во времени, отражая условия ее существования. Это хорошо иллюстрируют данные табл. 42 (по Pulliainen, Loisa, 1972). Процентное соотношение возрастных групп в отдельных популяциях было выяснено только для Лапландского и Печоро-Илычского заповедников (Семенов-Тян-Шанский, 1959), причем небольшое количество исследованного материала позволяет считать полученные данные самыми приблизительными. Было вычислено, что в первый год жизни из каждой тысячи глухарей гибнет: в Лапландском заповеднике — 520 и в Печоро-Илычском — 410 птиц (табл. 43). Все исследования возрастного состава глухарей на токовищах весной (Кириков, 1939, 1975; Семенов-Тян-Шанский, 1959) показали преобладание на них птиц в возрасте 2—3 лет. Среди глухарей, пойманных на току петлями между 2 и 5 мая, 19.4 % составляли годовалые птицы (Теплов, 1947а). Большой интерес представляют данные Киселева (1971),; основанные на результатах отлова птиц на галечниках (табл. 44), также фиксирующие более частое преобладание самцов.
Территориальность. На большей части ареала глухарь принадлежит к птицам вполне оседлым и проводит весь год в пределах сравнительно небольшой территории.
Запасы кормов, как летних, так и зимних на таких территориях, как правило, более чем достаточны, и птицам нет необходимости при сезонных изменениях питания предпринимать значительные перемещения.
Территориальная структура типичной оседлой микропопуляции выглядит следующим образом. Центром ее является токовище, т.е. место, куда птицы собираются для спаривания. В подавляющем большинстве случаев токовище отличается большим постоянством, хотя оно может «угаснуть», возникнуть или постепенно перемещаться год от года в каком-либо направлении.

Ряд наблюдений показывает, что при тесном соседстве самцы одной микропопуляции могут временами посещать токовище соседней, хотя документально такие случаи не доказаны. На токовище самцы имеют весной индивидуальные участки, величина и стабильность которых чрезвычайно изменчивы.
Остальное время года самцы и самки проводят в непосредственной близости от токовища, в радиусе до 3—4 км, где самки выводят птенцов, а самцы линяют.
Гнездовые территории самок не исследованы, хотя известно, что гнездятся они вблизи токовищ и не испытывают антагонизма друг к другу. Известны случаи, когда самки гнездились в непосредственной близости одна от другой. В Ленинградской обл. мне приходилось наблюдать регулярное гнездование отдельных самок примерно в одном и том же месте и известны случаи, когда в течение 2 лет самка гнездилась в одном и том же гнезде (Крутовская, 1958). Степень связи выводка с участком гнездования самки неизвестна, так же как неизвестны пространственные перемещения выводка в различные периоды развития. По моим наблюдениям, в Ленинградской и Новгородской областях Выводки по мере роста кочуют все более широко, но в целом в пределах радиуса в 1—2 км. Осенью, после распада выводков, птицы объединяются в стаи и всю зиму проводят в сравнительно однородных биотопах, но опять же в пределах территории данной микропопуляции.
Таким образом, связь отдельной особи с территорией у глухаря менее тесная, чем у рябчиков и дикуш. Индивидуальные участки у самцов появляются только на период спаривания, и они полностью разобщены с местами дневных кормежек и с местами жизнедеятельности в остальные сезоны. Несмотря на то что токовище вне времени размножения почти не посещается птицами, оно сохраняет свое значение территориального центра микропопуляции, члены которой не удаляются от него далее чем на 1.5—3.5 км.
Эту общую для вида картину подтверждают немногочисленные данные по кольцеванию птиц. Например, в Коми АССР из 14 возвратов 13 было получено от птиц, добытых или пойманных в 2—6 км от места кольцевания, и лишь один самец переместился за 15 дней на 40 км. В Кировской обл. из 30 случаев максимальное удаленна равнялось 3 км (Романов, 1960, 1979). Кольцевание в Финляндии показало, что большинство птиц, в особенности старые самцы, держатся в пределах радиуса 3 км от места мечения (один самец был добыт почти через 10 лет в 1 км от места кольцевания), Хотя отдельные птицы, в основном молодые самки, удалялись на 6—22 км.
Тем не менее в целом ряде мест, например по северной окраине ареала, а местами и на южной и даже кое-где в центральных его частях, связь микропопуляций глухарей с территорией далеко не так тесна. Это вызвано тем, что в местах гнездования нет либо достаточного запаса осенних или зимних кормов, либо мест, где можно добывать гастролиты. В результате с наступлением зимы птицы вынуждены перемещаться и такие перемещения нередко носят четко выраженный сезонный характер. У глухарей, как и у остальных тетеревиных птиц, даже во вполне оседлых популяциях существует склонность к перемещениям, которая особенно проявляется в осеннее время. В осенние месяцы странствующие одиночные глухари или же небольшие их группы могут появляться в самых неожиданных местах, нередко удаленных от обычных мест обитания на десятки километров. Так, по сообщению Теплова (1947а), в 20—30-е годы нашего столетия в октябре в Казанский зоопарк неоднократно поступали глухари, пойманные в городе. Птицы эти имели легкие травмы, возможно полученные при ударах о провода или строения, что может говорить о ночном или сумеречном времени перелета. Аналогичные сведения есть и для Москвы (Формозов, 1947), и для Ленинграда (данные автора).
Сезонные перемещения глухарей до сих пор почти не исследованы. На северной окраине ареала они, судя по собранным мною сведениям, происходят регулярно, принимая характер настоящих перелетов в годы с высокой численностью птиц. Северная граница ареала здесь подвижна и зимой отступает к югу. Так, на севере Западной Сибири глухари регулярно откочевывают на зиму из долинных елово-лиственничных лесов, расположенных севернее 66° с.ш. к югу по причине отсутствия здесь основного зимнего корма. То же самое происходит на западных и восточных склонах Приполярного Урала (Добринский, 1965). На западных склонах Приполярного Урала (65° с.ш.) такие перемещения мне удалось наблюдать в октябре—ноябре 1974 г. Из обширных пространств Печорской низменности, покрытых в основном болотами и узкими полосками еловых лесов, глухари перемещались на западные склоны Урала в поисках зимнего корма (хвоя и ягоды можжевельника, хвоя кедра) и гальки.
Во время таких перемещений численность птиц на горных склонах может возрастать в 10—15 раз, причем преобладают самки (соотношение полов 1 : 4; Естафьев и др., 1973). Эти перемещения могут, вероятно, продолжаться и далее к востоку. Во всяком случае на восточном склоне Урала, примерно на этой же широте (верховья р. Хулги) во второй половине октября наблюдался массовый перелет через реку в восточном направлении (Балахонов, 1978).
На обширных болотистых пространствах Западной Сибири осенние перемещения глухарей вызывает дефицит гальки, потребность в которой в это время резко возрастает. Обнажения с песком и гравием встречаются здесь почти исключительно по берегам некоторых рек, и перекочевки сюда глухарей носят массовый характер. Особенно известны благодаря истребительной деятельности браконьеров концентрации глухарей на галечных отмелях рр. Васюган, Тым, Кеть, Таз, Полуй, Надым, Пур. Концентрация птиц достигает здесь местами 150 и более птиц на 1000 га (Велижанин, 1968а). Первыми появляются старые самки, затем молодые, затем, через полмесяца, к ним присоединяются старые глухари и последними — молодые самцы-первогодки. Такие вылеты начинаются в первой половине сентября и достигают максимума в начале октября.
На отдельных отмелях в верхнем течении рек может скапливаться одновременно по нескольку десятков птиц (Назаров, Шубникова, 1975). Эти же авторы упоминают о случаях, когда в каких-либо определенных местах в обычное для «галькования» время глухарей почти не было, а затем они появлялись в довольно большом количестве (верховья рр. Полуй, Куноват, правый берег Оби).
Особый интерес представляет сообщение Соловьева (1927а) о сезонных перемещениях глухарей в северной части Печорского края. Он указывает, в частности, что осенью глухари покидают места гнездования в «парме» (глухие еловые леса с преимущественно моховым покровом) и перебираются в боровые районы, где сосна дает им основной зимний корм. Кроме этих передвижений отмечены также перемещения глухарей в долготном (с востока на запад) направлении, происходящие в конце марта—начале апреля. Эти перелеты происходят стайками по 20—50 птиц и, на мой взгляд, представляют собой обратное движение птиц на места гнездования. По наблюдениям охотников, такие перемещения происходят с определенной периодичностью, примерно 1 раз каждые 5—10 лет, что, по-видимому, также связано с возрастанием общей численности глухаря после удачных сезонов размножения. Д.К. Соловьев сообщает также о необычном осеннем перемещении глухарей по кольцевому маршруту у западных предгорий Северного Урала. Обобщив разрозненные сведения, он установил, что глухари начинают перемещаться в северном направлении примерно от долины р. Щутор и летят на север вплоть до границы леса и тундры (невдалеке от Усть-Усы). Далее птицы поворачивают вдоль края леса на запад и затем в районе с. Денисовки направляются обратно, уже в юго-восточном направлении, совершая таким образом кольцевой маршрут длиной около 700—1000 км. Во время этих перелетов птицы передвигаются стайками по 10—30 птиц и одиночками по ранним утрам. Они могут появляться в это время (начало октября) даже в поселках, где отдыхают на крышах. Во время такого перелета на р. Лебедь в 1908 г. два промышленника насчитали за час 500 птиц. Перелеты эти проходили с особой интенсивностью в 1923 г. и гораздо слабее. — в 1921 г.
Автор связывает эти перемещения с поисками гальки, упоминая случаи, относящиеся к 1921 г., когда в лесах по рр. Лыжа и Кожва находили зимой в лунках много замерзших глухарей, у которых в желудках не было гастролитов.
Впоследствии таких крупных перемещений никто не наблюдал, равно как и перелетов с одной стороны Урала на другую, о которых упоминал Мензбир (1895). Однако на Кольском п-ове О.И. Семенов-Тян-Шанский наблюдал иногда что-то похожее на перелеты: один раз (4.XI) стая самцов и самок летела высоко над лесом на северо-восток, а одиночные птицы наблюдались летящими над горной тундрой на такой высоте, что даже трудно было определить вид птицы.
По сообщениям местных охотников, летом 1959 г. в бассейне Турухана глухари летели стаями на довольно большой для них высоте (выше крон деревьев), самцы отдельно от самок, прямо на запад (Назаров, Шубникова, 1975). Эти данные говорят о существовании каких-то особых явлений в жизни северных популяций глухарей, которые нам пока неизвестны.
Регулярные сезонные перемещения глухарей происходят на западном Алтае, где птицы зимуют в кедрачах, отстоящих нередко на 50 км от мест гнездования. В частности, в конце марта—начале апреля Наблюдалось движение глухарей с кедрачей Ивановского хребта на хребет Листвягу и далее (Кузьмина, 1962). В Ульяновской обл. осенью глухарей встречали примерно за 50 км от ближайшего крупного леса (Житков, Бутурлин, 1906), а в Уфимской области «за 75—80 верст от их ближайшего обитания — Благовещенского завода» (Толстой, 1905). На Южном Урале, по наблюдениям Кирикова (1952), глухари совершают регулярные сезонные миграции, прилетая весной на места размножения в дубравы горного массива Шайтан-Тау и улетая на зимовку, видимо, к северу, в хвойные леса. Самих перелетов С.В. Кириков, правда, не наблюдал, и в своих выводах он базировался на появлении глухарей в Шайтан-Тау весной и исчезновении их осенью.
Образ жизни. Обыкновенный глухарь — типично лесная птица, но связь его с лесом не такая тесная, как, например, у рябчика. Время от времени глухарей можно видеть летящими высоко над лесом, самки во время вождения птенцов часто выходят на болота, на пойменные или субальпийские луга, а в местах контакта леса со степью глухари иногда выходят в степь для сбора насекомых, удаляясь от зарослей до 150 м (Сушкин, 1914). У верхней границы леса на Северном Урале глухари выходят кормиться на совершенно открытые ягодники (Теплов, 1947а), и я сам наблюдал, как они поднимаются в горную тундру и кормятся вороникой по соседству с тундряными куропатками. О.И. Семенов-Тян-Шанский тоже встречал молодых глухарей в открытой тундре вдалеке от леса.
Большую часть времени в году глухарь проводит на земле, несмотря на то что древесный корм является для него основным всю зиму. Наземный образ жизни особенно характерен для летнего времени, когда птицы очень неохотно взлетают и стараются уходить от опасности заблаговременно пешком, очень напоминая в этом отношении дикуш. В летний период глухари взлетают на деревья только на ночлег или в случае опасности. В горах вверх по склону птицы передвигаются пешком, вниз же под гору всегда слетают (Кузьмина, 1962).
Взлет с земли у самца шумный и тяжелый, но когда птица слетает с дерева или со склона, то шума почти не слышно. Самки способны к вертикальному взлету, что вряд ли доступно самцам, несмотря на сообщения об этом в литературе (Кузьмина, 1962); для этого самец слишком тяжел. Цри взлете с дерева птица сначала резко снижается, набирая скорость почти над самой землей. Заблаговременно уходящая от опасности самка взлетает практически бесшумно, делая сильный толчок ногами. Находясь в 20 м от взлетающей самки и специально прислушиваясь, можно уловить только легкий шум. В отличие от рябчика и дикуши обыкновенный глухарь при перемещениях нередко поднимается над пологом леса и может лететь на значительные расстояния. В зависимости от конкретных условий птицы летают на разных режимах, или делая равномерные и непрерывные взмахи крыльями (что характерно прежде всего для перелетов на короткие расстояния), или же чередуя несколько взмахов со скользящим планированием. Сколько может пролететь глухарь без остановки — неизвестно, но во всяком случае не менее 15—20 км. О.И. Семенов-Тян-Шанский неоднократно наблюдал глухарей, летящих высоко над лесом и даже над тундрой, причем птицы подолгу летели скользяще-планирующим полетом, не шевеля крыльями и со свистом рассекая воздух. По некоторым наблюдениям, скорость нормального полета глухаря — 60 км в час, а при преследовании хищником она достигает почти 96 км в час (Bannerman, 1963).
Стайность присуща глухарям в гораздо большей степени, чем рябчикам или дикушам. Собственно, глухари живут сообществами разной величины в течение почти всего года. В зимнее время на европейском Севере эти стаи в нетронутых лесных массивах достигают значительных размеров (до 20 и более птиц одного пола) (Теплов, 1947а; Seiskari, Koskimies, 1955). В Кировской обл., по данным Мензбира, осенние стаи самок достигали 100, а самцов — более 70 птиц, то же отмечено Семеновым-Тян-Шанским для Лапландского заповедника.
Нередко самцы и самки держатся в смешанных стаях. В южных районах ареала стаи эти значительно меньше по размерам, а в местах, где глухарей усиленно преследуют, птицы, в особенности самцы, нередко держатся одиночками. Весной глухари группируются на токах. И только в летнее время самки изолируются для вывода птенцов, самцы же проводят период линьки нередко небольшими группами по 2—6 птиц (Сушкин, 1938; Дулькейт, 1964), хотя в этот период одиночные самцы встречаются наиболее часто. Образование зимних стай происходит уже в сентябре. Видимо, как ток, так и зимняя стая объединяют птиц одной микрбпопуляции, но детальных наблюдений на этот счет нет.
Питание. Питание глухаря, в особенности зимнее, выглядит чрезвычайно специализированным даже на фоне глубокой специализации зимнего питания всех тетеревиных птиц. Основой его на большей части ареала в зимние месяцы являются хвоя сосны, обычно Pinus silvestris или Р. sibirica, с небольшой примесью молодых шишечек и кончиков веток. В теплые сезоны года, с мая по сентябрь, хвоя сосны также потребляется птицами, но в самом незначительном количестве. Есть, правда, указания, что во время линьки рамфотеки глухари хвою не едят.
Несмотря на резкую разницу в строении и величине челюстного аппарата у самцов и самок, существенной разницы в их питании подметить не удалось, в том числе и в питании сосновой хвоей. Специальные исследования на этот счет (Pulliainen, 1970а) показали, что небольшая разница заключается лишь в наличии в питании самцов шишечек сосны (0.4 % от всего содержимого зобов). Предпринятое мною исследование зобов 16 самцов и 7 самок, добытых в одной и той же местности зимой 1967/1968 г., показало также, что процентное содержание раздельно сорванных игл хвои у самцов составило 79.63 % всего содержимого, у самок — 83.08 %, отдельных же концевых побегов соответственно 19.97 и 16.92 %. Небольшое количество исследованного материала не позволяет считать эти различия статистически достоверными. По наблюдениям в Финляндии (Seiskari, 1962), самки в отличие от самцов объедают хвою с внутренних и нижних частей крон и веток деревьев, расположенных в глубине леса, и не выходят на открытые сосны, тогда как самцы кормятся на наружных и верхних частях кроны и предпочитают отдельно стоящие сосны на полянах или болотах. Самки, кроме того, несколько раньше переходят осенью на хвойную диету и раньше заканчивают ее весной.
В Финляндии же опытным путем было подтверждено явное предпочтение, оказываемое самцами хвое больных сосен (в частности, пораженных Peridermium sp.) и подсочных деревьев, поскольку в ней, как предполагается, из-за нарушений обмена веществ остается больше продуктов фотосинтеза (Seiskari, 1962). Химический анализ хвои, взятой из зобов птиц и с вершин кормовых деревьев, показал наличие в первой заметно меньше кальция и больше азота, что в общем-то характерно для однолетних игл; в 2—3-летних иглах кальция больше, а протеинов меньше (Pulliainen, 1970а). Предпочтение же самцами определенных деревьев, большей частью действительно угнетенных или пораженных болезнью, наблюдалось повсеместно и известно давно (Лобачев, Щербаков, 1936; Теплов, 1947а; Lindrot, Lingren, 1950; Кириков, 1952). На таких кормовых деревьях бывает объедено до 50 % хвои, а иногда и того более. Возможно, правда, что угнетенность таких деревьев — результат деятельности глухарей. Там, где обыкновенную сосну заменяет кедр, глухари столь же охотно едят хвою последнего, и нет никаких данных, говорящих о предпочтении одного из видов этих сосен.
Среди второстепенных кормов в зимнее время наибольшее значение имеет можжевельник (веточки с хвоей и плоды), а в Шотландии — хвоя псевдотсуги (Zwickel, 1966). В тех же случаях, когда глухари зимуют в лесах, лишенных сосны или же имеющих ее в ничтожном числе, основу зимнего рациона в зависимости от обстоятельств может составлять хвоя пихты (Удмуртская АССР, Теплов, 1947а; Алтай, Дулькейт, 1964), хвоя ели (Западный Алтай, Долгушин и др., 1962; южные части Кольского п-ова, Семенов-Тян-Шанский, 1959; Карпаты, Островский, 1974), почки и концевые побеги березы (также и сережки), дуба, осины, липы, ольхи, вяза, ивы (Рязанская обл., Мензбир, 1902; Южный Урал, Кириков, 1952; Северный Урал, мои данные). В отдельных случаях сережки березы составляют до 15 % от содержимого зоба (Новосибирская обл.; Телепнев, 1972). Наконец, в горах Кантабрии основной зимний корм составляют жесткие листья вечнозеленого надуба (Kastroviejo, устное сообщ.). Эти факты хорошо иллюстрируют способность при столь высокой в общем-то стенофагии в зимнее время менять основной корм. В обычных же условиях зимой глухарям нужна именно сосновая хвоя. Например, в Ленинградской и Новгородской областях, как и во многих других местах, в начале зимы стебли черники нередко остаются неприкрытыми снегом вплоть до конца декабря, но все равно глухари используют этот столь любимый осенью корм только как небольшую добавку к исключительно хвойному рациону.
Весенние, летние и осенние корма гораздо более разнообразны и в сильной степени зависят от характера и богатства растительности данной местности.
Однако и здесь выделяются несколько видов растений, имеющих первостепенное кормовое значение. Это прежде всего черника — ее побеги и листья глухари поедают, как только они покажутся из-под снега, летом же и вплоть до поздней осени в корм идут завязи, зеленые и спелые ягоды черники (поздней осенью черника является только добавочным кормом к хвойному рациону). Такое большое значение черника имеет на большей части ареала, за исключением только самых южных его частей. В меньшем числе, но также регулярно используются и другие виды рода Yaccinium, а также ягоды вороники (Empetrum nigrum). Весной птицы, особенно самки, охотно едят свежую зелень, цветки и соцветия трав и древесных пород. Летом в рацион включаются семена различных трав (осоки, марьянника), и что особенно характерно, листья деревьев и кустарников — ив, осины, березы, ольхи, а также различные хвощи и папоротники. Летом и в начале осени употребляется и животная пища — слизни, пауки, различные насекомые, среди которых предпочитаются кузнечики, муравьи и личинки пилильщиков. Удельный вес животных кормов в питании глухаря невелик, не выше 10 %; особенно охотно насекомых поедают линяющие птицы (Теплов, 1947а).
Осенью основным кормом становятся ягоды. В ряде районов большое значение приобретает хвоя лиственницы, листья осины, кедровые орехи и желуди, а такжё злаковые культуры. Осеннее питание глухарей в той или иной части ареала может быть весьма различным.
Поедание гальки для глухарей очень важно, потребность в ней может явиться причиной уже упоминавшихся массовых перемещений птиц. Количество гальки в желудках может достигать 71 г у самцов и 27 г у самок. Активные поиски гальки глухари предпринимают и зимой, и весной с началом таяния снега и особенно осенью. Летом нужда в гальке, видимо, наименьшая, и в августе в желудках птиц количество ее минимально. Наиболее интенсивные поиски и поглощение гальки происходя? с середины сентября, в особенности после первых заморозков, когда глухари начинают переходить на зимний корм. На утренних зорях глухари вылетают в поисках ее на берега рек, грунтовые дороги, косы, отмели, песчаные карьеры и обрывы террас. В Мещерской низменности первое появление глухарей «на гальке» отмечено в начале августа (Киселев, 1971).
В течение зимы количество гастролитов в желудках глухарей постепенно уменьшается, но все же они сохраняются до весны. По-видимому, в течение зимы глухари от случая к случаю пополняют свой запас гальки. Есть данные о том, что глухари выкапывают гальку из-под снега (Назаров, Шубникова, 1975).
В северной тайге глухари употребляют камешки диаметром от 2 до 5 мм (средний диаметр камешков у самцов 3.5, у самок 3.2 мм). Их количество у самцов осенью, может достигать в среднем 900 штук, при общей массе 46.4 г, у самок — 29.9 г. Содержание гастролитов в желудке к весне падает и во время токования масса их в среднем — 33.2 г (Романов, 1975).
В ряде мест, где добывание гальки зимой затруднено, она заменяется косточками ягод шиповника и черемухи, причем степень упитанности птиц явно не зависит от сорта гастролитов в их желудках (Новосибирская и Томская области; Телепнев, 1972). Весенние вылеты на гальку изучены заметно меньше вследствие меньшей их интенсивности и продолжительности. Как и осенью, они особенно хорошо выражены там, где отсутствуют обнажения и рельеф в основном равнинный. В Мещерской низменности длительность весенних вылетов на галечники прямо зависит от характера весны, продолжаясь всего 12 дней в дружные теплые весны и вдвое дольше при затяжной и холодной весне. Наиболее яркой особенностью весеннего «галькования» является заметно большая потребность в гальке у самок, которые составляют 70—80 % вылетающих на галечники птиц. Весь период «галькования» совпадает с разгаром токования. Это свидетельствует о резком возрастании потребности самок в гальке, что явно связано с откладкой яиц (Киселев, 1971).
О потребности глухарей в воде известно немного. Теплов (1947а) сообщает, что пьют они регулярно и не менее чем дважды в сутки, утром и вечером, а на Южном Урале в засуху как выводки, так и старые птицы концентрируются у источников (Юргенсон, 1939; Кириков, 1952). По наблюдениям за птицами в неволе, глухарята склевывали капли воды с растений уже в первые дни жизни, но некоторые птенцы совершенно не пили воду в течение 25—60 дней (Крутовская, Крутовская, 1953). Зимой глухари, как и другие тетеревиные, поедают снег во время ночевок в лунках.
Зимний сезон. Зимнее время глухари проводят так.же, как и остальные тетеревиные. Территориально они в это время связаны с сосняками, причем в последних выбирают места с минимальной кухтой, стараясь кормиться хвоей, свободной от снега и льда. Зимой держатся у опушек, в разреженных насаждениях, у гребней хребтов, где снег быстрее сбивается с деревьев ветром. Группы или стаи самцов и самок подолгу, иногда всю зиму, держатся около облюбованных кормовых деревьев, и район обитания такой группы может колебаться от 0.2 до 50 га (Теплов, 1947а; Ивантер, 1965; мои данные). Обеспеченность кормом у глухарей в сосняках настолько полная, что они действительно могут зимовать на небольшом «пятачке», питаясь на окружающих деревьях. Потребность в хвое у глухарей довольно высока. Суточный рацион, по различным определениям, 440—550 г хвои в день (Лобачев, Щербаков, 1936; Теплов, 1947а; Семенов-Тян-Шанский, 1959). По данным О.И. Семенова-Тян-Шанского, максимальное количество хвои, обнаруженное в зобах у Кольских глухарей, отличающихся мелкими размерами, равнялось у самцов 242 г, у самок 148 г, или 8.3 % от общей массы птицы. Максимальное количество съеденной за день хвои при содержании в вольерах равнялось у самца 375 г, минимальное 20 г, у самки соответственно 150 и 0 г (Seiskari, 1962). Мы можем принимать близкими к дневному рациону последние максимальные показатели, учитывая, что глухари содержались на смешанном корме и хвоя была не единственным источником питания. Старый самец, только что забравшийся в снежную камеру (в 17 ч) и добытый А.В. Андреевым в начале февраля, имел в зобу 374 г сосновой хвои, что составляло 7.5 % от массы птицы. По наблюдениям С.П. Кирпичева в вольерах, количество хвои, потребляемое за день птицей, зависит от температуры воздуха и, чем ниже последняя, тем больше съедает птица. По моим расчетам, среднесуточный рацион самца зимой составляет 282.35 г хвои (сухая масса).
Длительность и количество кормежек глухарей зимой определенно зависят от температуры воздуха и длины светового дня. На Кольском п-ове, на 67—68° с.ш., где минимальный световой день длится 6 ч, птицы кормятся дважды в день — на рассвете и затем вечером. В случае морозов середину дня, как и ночь, птицы проводят под снегом. При вольерном содержании в Финляндии глухари кормились зимой все светлое время суток, причем кормовую активность стимулировали высокое давление и низкая температура воздуха (эти два явления в указанных местах достаточно очевидцо связаны между собой). По моим наблюдениям в Ленинградской обл., в морозы —30 °С глухари кормятся все светлое время суток, без перерыва в середине дня. По данным Теплова для Печоро-Илычского заповедника, у глухарей в период декабрь—февраль бывает только одна кормежка около полудня и в морозные дни она длится не более полутора часов.
Все остальное время йроводится в снегу. В случае же морозов ниже —30°С или же сильных метелей и северных ветров глухари вообще не выходят на кормежку, оставаясь под снегом по 3—4 сут. Об одной кормежке в день у глухаря на Алтае при сильных морозах пишет и Дулькейт, температурным «порогом», после которого птицы не выходят на кормежку, является там температура —35°С. У О.И. Семенова-Тян-Шанского, однако, приводятся конкретные наблюдения за кормежками глухарей при —33 и —36.5 °С. При этом, правда, глухари избегали взлетать на деревья, а бродили по снегу, склевывая побеги можжевельника, или взлетали на низкие сосёнки, переходя от одной к другой пешком. Следует учесть, что сведения о длительном, более суток, пребывании глухарей в подснежных камерах не подтверждены детальными наблюдениями.
Различные типы поведения глухарей в середине зимы хорошо иллюстрируют следующие наблюдения, проведенные мною в Ленинградской обл.
21 XII 1970. Ясно, безветрие, атмосферная температура —21°С. 16.00. 5 самцов прилетели на ночевку с окраины высокоствольного леса в низкий и разреженный сосняк на окраине верхового болота. Опускаясь в рыхлый, накануне выпавший снег, тормозили хвостом, затем в соприкосновение со снегом приходили лапы и, наконец, туловище. Длина тормозной полосы на снегу — 5 м, из них первые 332 см — след полураскрытого хвоста, затем идут борозды от лап и в конце ее — ямка от туловища. Двое из пяти сели более круто, не оставив тормозной полосы, и стали зарываться в снег сразу. Остальные трое прошли каждый 1—1.5 м по снегу, проваливаясь на 8—10 см, и тоже стали закапываться. Один из самцов проделал метровой длины тоннель, прежде нем устроил нонлежную камеру, остальные делали только короткий вход длиной 40 см. Вся эта группа накануне ночевала здесь же, лунки вчерашней ночевки в 10 м.
22.XII.1970. Утро ясное, безветрие, атмосферная температура —26 °С. Птицы стали покидать камеры в 8 ч 20 мин—8 ч 30 мин (восход солнца — 8 ч 30 мин). Сначала высовывали головы наружу, оглядывались, затем спокойно выходили на поверхность, разламывая кровлю.
Трое, покидая камеры, оставили там экскременты слепых кишок, один сделал это спустя 10 мин, уже начав кормежку, а один, пройдя 11 м, снова лег на поверхность снега, пролежал здесь минут 7 и затем оставил здесь экскременты. К 9 ч утра все глухари дружно кормились, расхаживая по снегу и объедая короткую желтоватую хвою на низких, до 2.5 м высоты, сосенках. Объедая хвою с нижних ветвей, глухарь обходил сосенку вокруг, вытаптывая кольцо следовой канавки, так что дерево как бы оказывалось на своеобразной снежной клумбе.
Время от времени то один, то другой самец взлетал на ‘верхушки сосенок, кормился около 15 мин на верхушках и затем снова соскакивал вниз. Эта деятельность продолжалась до 13 ч 20 мин, когда птицы одна за другой стали устраиваться на солнцепеке в поверхностных лупках. При этом они топорщили оперение, как бы отряхиваясь, оправляли его клювом и затем впадали в полудремотное состояние, время от времени оглядываясь. Перед тем как залечь, примерно в 13 ч, один из самцов, перестав кормиться, вдруг поднял и раскрыл хвост и принял токовую позу, затем пошел по снегу, чертя его слегка отставленными и приспущенными крыльями. Затем он проделал токовой взлет, прошел еще немного в токовой позе, постоял немного, оглядываясь, и затем, сложив слегка взъерошенное оперение и хвост, вновь принялся за кормежку. Остальные самцы не обратили на его маневры никакого внимания и спокойно продолжали кормиться. В поверхностных лунках глухари находились около часа (от 40 до 67 мин), после чего возобновили кормежку, но теперь они больше времени тратили на переходы от дерева к дереву, а двое из них опять залегали минут на 20. Птицы стали часто и подолгу осматриваться и в 16 ч 10 мин вдруг дружно взлетели и перелетели примерно на 300 м; усевшись на верхушках сосен в кромке леса, они оставались здесь до заката, слетев далее на ночевку в сторону болота. Покидая место кормежки, глухари взлетали полого, оставляя при этом на снегу 4—6 ударов крыльями. Расстояние между первыми 6 взмахами у одного из них оказалось 88, 128, 110, 135, 130, 125 см.
Другое наблюдение на том же болоте.
14 II 1976. Ясно, легкий ветер, атмосферная температура —16°С. 4 взрослых самца накануне устроились на ночевку среди редких болотных сосенок в 400 м от кромки леса. Глубина снега — 52 см. Примечательно, что все четверо устроили открытые лунки — снежные ямки глубиной 26—28 см, без кровли, хотя глубина снега и его рыхлость вполне позволяли устроить подснежную камеру. Один из глухарей в середине ночи (было полнолуние) покинул свою лунку, прошел 50 см и снова закопался, но уже поглубже, хотя опять же без крыши. Остальные тоже примерно в это же время сменили место ночлега, переместившись на 80—120 см и устроившись в таких же по глубине лунках. Примерно в 8 ч 30 мин птицы стали покидать-лунки, причем трое сразу же взлетели на небольшие сосенки, где и оставили экскременты слепых кишок. Только после этого, оглядевшись и поклевав немного хвою, глухари слетели на снег и начали обычную пешую кормежку, поедая хвою с низких сосенок и утаптывая вокруг них снег. Четвертый же, менявший ночью лунку на более глубокую, оставил ее несколько позже других, прошел около 10 м, склевывая хвою с сосенок, стоявших на дороге, постоял с минуту, а затем принял токовую позу и совершил токовой взлет, пролетев 2 м. Затем он выполнил два взлета подряд, один — со сменой направления, побудив и остальных к тому же. Все самцы начали проделывать токовые взлеты в разных направлениях, при посадках оставляя на снегу отпечатки полураскрытого хвоста (ширина — 39 см).
Каждая из птиц проделала по 2—3 взлета, а один из самцов принял после такого взлет «распластанную позу» (см. далее), дав мне возможность измерить по отпечаткам на снегу размах его крыльев (155 см). После этого короткого оживления кормежка продолжалась обычным образом до 16 ч 20 мин. Самцы объедали хвою, иногда срезая ее концевые пучки вместе с кончиком веточки. В средине дня они примерно полтора часа отдыхали в мелких поверхностных лунках. В 16 ч 20 мин они взлетели на сосенки, посидели 15 мин, почти не кормясь, и затем полетели к кромке леса, где вновь их найти в тот день не удалось.
В зимнее время глухари бродят по снегу довольно часто, но глубокий рыхлый снег все же существенно затрудняет их передвижение. Благодаря роговой бахроме и длинному оперению плюсны площадь поверхности следа глухаря увеличивается примерно вдвое против летней (52 см3 против 26 см2) и нагрузка с 80 г/см2 падает до 40 г/см2 (Теплов, 1947а; Дулькейт, 1964). По моим расчетам, площадь обеих лап самцов составляет 70.2—86.4 см2, а нагрузка на след, вычисленная у конкретных птиц с известной массой, составляет 44.5—53.8 г/см2.
Все же в рыхлом снегу глухарь сильно вязнет; по’наблюдениям Дулькейта, он может проваливаться на глубину 11—18 см и оставляет за собой борозду шириной до 21 см. При таком состоянии снегового покрова глухари редко проходят большие расстояния: в одном случае на 50 м пути самки было насчитано 6 поверхностных лунок для отдыха.
Как и остальные северные тетеревиные, глухари на большей части ареала проводят длинную зимнюю ночь, а нередко и большую часть дня под снегом, как только позволит глубина снегового покрова. По наблюдениям Дулькейта на Алтае, минимальная глубина, годная для устройства лунки, — 40 см, Семенов-Тян-Шанский указывает минимальную глубину снега для самки 40 см и для самца 50 см. В начале зимы, когда снега еще мало, глухари выискивают места, где снега больше, т.е. птица явно стремится как можно скорее перейти на ночевку под снегом. Именно в конце осени, когда бывает очень мало снега, глухари должны более всего страдать от холода. В частности, гибель глухарей от истощения во время сильных морозов и при невозможности зарывания в снег наблюдалась при вольерном разведении (Немцов и др., 1973). Поэтому к началу зимы у глухарей северных популяций накапливаются значительные жировые отложения (Романов, 1975).
Глухари погружаются в снег так же, как и остальные тетеревиные, либо зарываясь тотчас по приземлении, не оставляя следов, либо же пройдя некоторое расстояние в поисках наиболее подходящего места. В отдельных случаях как самцы, так и самки прежде, чем устроить камеру, проходят под снегом расстояние до 3 м, временами высовывая голову для обзора. Толщина кровли не бывает больше 30 см, в среднем 15—20 см (Теплов, 1947а; Дулькейт, 1964).
По наблюдениям последнего на Алтае, глухари в некоторых случаях в течение ночи меняют место ночлега: покидая старую камеру, птица продвигается под снегом дальше на 1.5 м и отрывает вторую камеру, где проводит остаток ночи.
Мне приходилось наблюдать нечто подобное в Ленинградской обл. в марте (см. выше). Следует заметить, что ходы под снегом глухари делают при глубине снежного покрова не менее 40 см.
С увеличением светового дня и при небольших морозах глухари кормятся утром и к вечеру, а середину дня нередко проводят в поверхностных лунках, на солнце. При температуре воздуха около 0 °С глухари, как и остальные тетеревиные, под снегом не ночуют, а спят, забившись в кроны елей и сосен, либо же под нависающими лапами елок на поверхности снега. Именно этим обстоятельством объясняется тот факт, что южная популяция глухарей в Финляндии в отличие, например, от северной мало ночует в снегу (Seiskari, Koskimies, 1955). Ближе к весне, с появлением наста глухарь гораздо больше времени проводит на поверхности снега, много ходит и кормится здесь же, не взлетая на деревья; ночевки на поверхности снега, а не в подснежных камерах становятся регулярными! О.И. Семенов-Тян-Шанский указывает, что при крепком насте глухари и не пытаются пробивать его и спят на поверхности иногда под елью, а иногда и открыто.
Бюджет времени в середине зимы при температурах -10 -30 °С на широте 60° (Ленинградская обл.) по большому числу наблюдений выглядит следующим образом: пребывание в подснежной камере (сон) — 16.41 ч, пребывание в поверхностной лунке (обычно дневной отдых) — 2.42 ч, кормежка с хождением по снегу и взлетами на невысокие сосны — 4.88 —5.074 ч, полет — 0.1 ч.
Таким образом, для зимы характерно резкое снижение двигательной активности. Потери массы за зиму составляют на европейском Севере у самца 3.33 и у самки 1.46 г в сутки. Суточная продукция экскрементов составляет у самцов: твердых экскрементов — 150 г, жидких — 34 г.
Помет обыкновенного глухаря легко узнается как благодаря большому размеру, так и консистенции, поскольку содержит в основном перемолотую и спрессованную хвою. Длина твердых экскрементов у самца от 42 до 58 мм, диаметр 11—13 мм, масса от 1.24 до 1.47 г. В среднем зимний бюджет энергии составляет у самца в сутки 5735.1 кДж, из него энергия существования составляет 1558.3 кДж/сутки, т.е. 27.1 % от всей энергии и экскреторная энергия — 4185.2 кДж/сут (3246.9 кДж выделяется с твердыми и 938.3 кДж — с жидкими экскрементами). Следует учитывать, что это — усредненные данные. Все величины, составляющие суточный бюджет энергии, могут заметно меняться день ото дня, находясь в тесной зависимости от физиологического состояния птицы, погодных и кормовых условий. Примечательно, что канадская дикуша, питаясь сосновой хвоей, усваивает 29 % энергии, в ней заключающейся (Pendergast, Boag, 1971b).
Размножение. Обыкновенный глухарь — типичный полигам с групповым характером тока. Процесс токования и все, что с ним связано — место токования (токовище), пространственная и популяционная структура тока, сроки — составляет довольно сложную систему, которая обеспечивает преимущественное участие в размножении наиболее жизнеспособных самцов, прошедших испытание естественным отбором в первые 2 года жизни. Это обеспечивается прежде всего иерархической структурой тока и свободным выбором самцов самками.
Самцы достигают половозрелости, как и самки, в течение первого года жизни. При вольерном разведении содержащиеся поодиночке молодые самцы активно токуют уже в первую весну и активно, спариваются, однако процент оплодотворенных яиц в таких случаях очень низок (Кутовая, 1976), и, по-видимому, жизнеспособность птенцов тоже низка. Очень показательно, что при групповом токовании самцов в вольерах в группе токует только «лидер», остальные молчат. В природных условиях годовалые самцы при наличии доминирующих самцов более старших возрастных групп практически не имеют никаких шансов на спаривание, поскольку, будучи подавлены токующими старыми самцами, не могут даже частично выполнять брачный ритуал и, по моим наблюдениям, полностью игнорируются самками (Потапов, 1975). Однако при полном истреблении на току взрослых самцов центральные участки занимались молодыми птицами, которые токовали и размножались (Кириков, 1952). По данным Романова (1980), в нормальных условиях самцы занимают участки на току в возрасте около 2 лет, но в это время они еще не отличаются большой активностью. Самки также вступают в размножение на следующую весну далеко не всегда. Данные, полученные как в природе, так и в вольерах, показывают, что на первом году жизни вступают в размножение только те самки, масса которых ко времени спаривания не меньше 1800 г (Киселев, 1971; Немцев и др., 1973). Масса же самок зависит как от условий зимовки и ранней весны, так и от времени рождения — самки из поздних выводков не успевают набрать нормальной массы к зиме.
Весь процесс токования происходит на специальных участках — токовищах. Места расположения токовищ, их размеры и характер растительности на них довольно разнообразны, однако же есть некоторые особенности, которые сохраняются в любой области. Прежде всего токовище располагается под пологом леса и никогда не устраивается на открытых местах. С другой стороны, лес на токовище не бывает очень густым, подрост и подлесок развиты незначительно — лес на токовище просматривается во все стороны по крайней мере на 50 м. На большей части ареала, особенно в равнинных местах таежной зоны, токовища располагаются в участках соснового или смешанного леса, большей частью заболоченного, прилегающего к массивам верховых сфагновых болот.
Это могут быть перелески между болотами или вдающийся в болото лесистый мыс, причем токовища часто располагаются на более возвышенных участках (гривах, сельгах и т.п.). Токовище может находиться и среди сплошного леса, но в таких случаях оно обычно приурочено к участкам заболоченного сосняка с более разреженным древостоем и низкими деревьями, либо же, в более сухих местностях, у границ высокоствольного леса с мелколесьем. Характер леса на токовище варьирует незначительно. Обычно, преобладает сосна, нередко с примесью березы, ели и ольхи, но бывает, что преобладают береза или ель.
Почва большей частью чистая, не захламлена валежником, но иногда бывает густо заросшей невысоким вереском. Часто во время токования почва на токовище почти сплошь залита водой либо же представляет пропитанный водой мох. В южной части ареала тока располагаются и в лиственном лесу (дубняки, осинники, березняки и т.п.). В горах токовища размещаются большей частью у верхней границы леса: на Алтае, например, в высокоствольных кедрачах и сосняках в верхней части склонов или на гребнях и перевальных плоскостях, где лес не захламлен, подлесок разрежен, а зимой из-за постоянных ветров деревья свободны от снега (Дулькейт, 1964). В ряде областей токовища располагаются в ельниках (западный Алтай, Кольский п-ов, Альпы, Карпаты, как исключение — в Ленинградской и Кировской областях).
Размеры токовища также весьма различны и зависят от численности птиц, от интенсивности охоты на токах и от характера местности. Там, где численность птиц высока, а специальной охоты на токах нет, токовища могут быть очень большими и тянуться на километры. На подобных токовищах собирается до сотни и более самцов. В недалеком прошлом такие токовища были известны для Белоруссии, Кировской обл., Коми АССР. Тока по 50—70 самцов существовали в Мордовской АССР, в Тюменской и Курганской областях, на северо-восточном Алтае. Площадь крупных старых токовищ в Печоро-Илычском заповеднике достигала 1.5 км2, причем у популяции, объединенной одним токовищем, бывают еще 2—3 второстепенных токовища, которые могут использоваться самцами поочередно,: крупные тока объединяют птиц на территории радиусом 2.3—3 км (Теплов, 1947а). На Алтае некоторые токовища тянутся по гребням гор на километры (Дулькейт, 1964). В местностях с сильным «охотничьим прессом» размеры токовищ и количество собирающихся на них птиц бывают, как правило, много меньше, причем во многих районах размеры токов за последние десятилетия продолжают неуклонно уменьшаться. Например, на лучших токах Ленинградской обл. еще недавно, в 60-х годах, собиралось до 20 —25 самцов, сейчас же такие тока практически исчезли. В настоящее время в Западной и Центральной Европе, в южной части лесной зоны Восточной Европы преобладают тока, собирающие по 2 —5 самцов, причем нередкими стали случаи, когда самцы токуют в одиночку. Одиночное токование для глухаря — явно вторичное явление, в целом виду не свойственное и обычно предшествует полному исчезновению птиц из данной области.
Средний размер небольшого токовища не превышает в поперечнике 300 м, занимая площадь до 10 га. В южной Финляндии в районах с нормальной численностью глухарей площадь токовищ колеблется от 6 до 20 га и число токующих на них самцов — от 6 до 18 (Pirkola, Koivisto, 1970). Площадь одного из токовищ под Ленинградом, на которое слеталось 7 самцов, была 300X 200 м, площадь же другого, где собиралось до 17 самцов, составляла 30 га (ЮООх 300 м).
В той же Ленинградской обл. тока в подходящих местах располагаются в 3 — 4 км один от другого (Пукинский, Роо, 1966), т. е. «радиус действия» тока составляет 1.5—2 км. Примерно такие же расстояния между токами отмечены и в Мордовском заповеднике — 2—5 км (Щербаков, 1967).
Места токов отличаются большим постоянством и используются многие десятилетия подряд. Птицы не бросают токовищ, несмотря на интенсивную охоту, рубки, появление в непосредственной близости человеческих поселений.
Только сплошная застройка, вырубка или поголовное истребление токующих птиц может уничтожить ток. Например, в Мордовском заповеднике до сих пор существуют тока, известные задолго до 1917 г. В то же время за последние 20 лет там же наблюдалось возникновение новых токов, а кроме того, стали характерными небольшие тока с коротким сроком существования, от 1 до 3 лет. Возможно, что здесь мы имеем дело с вспомогательными, «запасными» токовищами, но вопрос этот может решить только работа с мечеными птицами. Многолетние наблюдения в Мордовском заповеднике показали, что степень постоянства токовищ неодинакова. В одном случае токовище оказалось совершенно неизменным в течение 6 лет, причем даже индивидуальные участки самцов не изменились.
В других же случаях наблюдались перемещения токовищ на несколько десятков или даже сотню метров в течение нескольких лет. Максимальные по дальности перемещения не превышали 1.5—2 км (Щербаков, 1967). В Ленинградской обл. в одном случае токовище переместилось за 20 лёт на 1 км, в другом же за тот же срок — на 500 м. Известны лучаи, когда при зарастании токовища густым подлеском глухари переставали его посещать. Остается открытым вопрос о взаимоотношениях птиц, токующих и спаривающихся на соседних токовищах, расположенных близко друг к другу. Как мои наблюдения, так и впечатления других авторов (Теплов, 1947а) заставляют предполагать, что некоторые самцы, а возможно, и самки регулярно посещают не одно, а два и даже более токовищ. В. П. Теплов предположил даже наличие у микропопуляции «запасных» токовищ. Однако немногочисленные исследования, проведенные с мечеными птицами, показывают все же четкую привязанность самцов к определенному токовищу (Островский, 1973; Романов, 1979).
Жизнь микропопуляции глухарей тесно связана с токовищем не только в период токования. Самки устраивают свои гнезда в непосредственной близости от него, поблизости проводят лето и зиму самцы и молодые птицы. Зимняя стая глухарей обычно объединяет птиц, посещающих одно и то же токовище.
Сказанное не относится, конечно, к тем случаям, когда зимние и летние местообитания птиц находятся на значительных расстояниях, как, например, на Южном Урале или на северной границе ареала.
Сам процесс токования отличается большой сложностью и разнообразием, находясь в зависимости от многих факторов, среди которых численный состав популяции, погодные условия и степень воздействия со стороны хищников и человека — одни из главных. Большое разнообразие в процесс токования вносят индивидуальные особенности птиц, которые в рамках видового стереотипа поведения демонстрируют массу вариантов различных этологических реакций. В этом отношении обыкновенный глухарь заметно отличается от остальных тетеревиных птиц, брачное поведение которых более стандартно.
Процесс токования отчетливо делится на три периода или фазы. Первый период — подготовительный, или начальный, — охватывает время от самых первых проявлений токового возбуждения самцов в марте до их концентрации на токовищах и появлении на последних самок.
Второй период — основной. Это время, в течение которого самки регулярно посещают ток. Последний период — угасание тока, происходящее после окончания спаривания. Периоды эти выражены вполне четко, для каждого из них характерны определенные особенности в поведении как самцов, так и самок, и варьировать могут только календарные сроки и длительность самих периодов, в особенности начального и конечного. Находясь под фотопериодическим контролем, эти периоды в определенных пределах корректируются ходом весны.
Весеннее оживление, начинается ранней весной, в марте, а иногда, особенно в южной части ареала, и в феврале. Большей частью его наступление провоцирует ясная солнечная погода, сопровождаемая образованием наста. В такие дни во время кормежки отдельные самцы могут принимать токовые позы и даже совершать токовые взлеты. Расхаживая в основной токовой позе, самец оставляет на снегу следы от приспущенных крыльев, именуемые в охотничьей практике «чертежами» или «набродами». Такие элементы токового поведения птицы могут демонстрировать как на местах кормежек, так и на токовищах — четкой кривязанности к токовищу у самцов в это время еще нет. Бывает, что в группе кормящихся самцов спонтанно возникает «весеннее настроение», они начинают провоцировать один другого на различные токовые действия и в результате птиц настолько охватывает весеннее возбуждение, что они начинают гоняться друг за другом, прыгать, совершать токовые взлеты. После этого место, где кормились глухари, по следам на снегу напоминает токовище в разгар токования. Такой «ложный» ток обычно никогда не привязан к токовищу и может произойти в любом месте, где кормятся птицы. Одновременно с усилением брачной активности самцов начинается постепенное передвижение глухарей к токовищам. Характер этих перемещений имеет значительные популяционные различия: они мало заметны у оседлых популяций и принимают характер настоящих перелетов там, где места зимовок и размножения удалены друг от друга на значительные расстояния.
С усилением токовой активности самцы все чаще навещают токовища. Они начинают ночевать на нем, и утром, после пробуждения, в случае хорошей погоды некоторое время токуют, после чего уже приступают к кормежке. Чем больше самцов оказывается в это время на токовище, тем возбужденнее они себя ведут. В это время самцы территориально еще не связаны со своими токовыми участками и перемещаются довольно свободно во всех направлениях.
Только иногда отдельные самцы выбирают себе небольшие участки — «точки», где топчутся по утрам, подпрыгивают и выбивают в снегу округлые площадки с углублением посередине. Временами такое утреннее токование идет на глазах у самок, кормящихся в кронах окружающих деревьев и не проявляющих никакой заинтересованности в происходящем. Песни в это время исполняются еще редко. Самцы иногда начинают угрожать друг другу, но до драк дело еще, как правило, не доходит. Правда, иногда схватки все-таки происходят, о чем свидетельствуют вытоптанные в снегу площадки со следами крови и многочисленными шейными перьями, утерянными во время драки.
Все это начальное время токование идет нерегулярно, недолго, нередко вне основного токовища, и любое ухудшение погоды его прерывает. По данным Кирикова (1947, 1952), первыми начинают токовать самцы в возрасте старше 2 лет, затем уже 2-летние и потом годовалые самцы. Постепенно, с повышением температуры воздуха и таянием снега ток становится все более интенсивным и концентрируется исключительно в пределах токовища, хотя по-прежнему интенсивность токования сильно зависит от погоды. Эта зависимость вообще сохраняется до самого конца токования, но становится минимальной в разгар тока. На интенсивность тока особенно отрицательно влияет сильный ветер, с осадками или без них. Видимо, сильный шум в лесу при такой погоде делает токование далеко небезопасным для птиц. Примечательно, что в горах глухари избирают места для токования там, куда не доходит шум горных потоков.
Столь же отрицательно действуют осадки в виде снега и дождя, хотя в отдельных случаях глухари азартно поют и во время проливного дождя. Неоднократно замечалось также, что глухари предчувствуют погоду и перед ее ухудшением плохо токуют или же, наоборот, активно токуют в плохую погоду перед ее улучшением.
Второй, основной период токования характеризуется четко выраженным территориализмом самцов, их высокой активностью и агрессивностью. Самки регулярно появляются на току перед рассветом, посещая токовые участки отдельных самцов для спаривания. Этот период длится недолго, около 2 недель.
Особенно интенсивно протекает вторая фаза этого периода, когда происходит большая часть спариваний, причем длительность этой фазы максимальной активности обычно не превышает 7—10 дней (Теплов, 1947а; Pirkola, Koivisto, 1970). Этот период имеет наиболее твердые сроки, но и здесь погодные условия могут несколько растянуть его или, наоборот, сжать (в пределах 2—3 дней).
В средней части ареала период этот падает обычно на время появления и быстрого роста проталин, но в северной части ареала в разгар спаривания часто еще лежит сплошной снежный покров, хотя уже и идет его бурное таяние (Карелия, 66° с.ш., Потапов, 1975). Это объясняется тем, что через 3—4 дня после первого спаривания самка приступает к откладке яиц, для чего необходимо наличие свободных от снега мест. При отсутствии таковых в отдельных случаях самка может начать кладку яиц прямо на снегу (см. далее). В тех местах, где глухари зимуют на значительном удалении от мест размножения, активный ток начинается сразу же по прилете на место, а предварительные стадии проходят на зимовках (детали этого явления не изучены). Самки, появляясь на токовище в предутренних сумерках, в первое время летают с призывным криком в разных направлениях и садятся на землю по две, по три в любых местах, но постепенно у отдельных самок устанавливается связь с определенными самцами и впоследствии, в разгар спаривания, они прилетают прямо на их токовые участки. Общая схема токования в это время такова.
Во время дневных кормежек самцы не удаляются далеко от токовища и появляются на нем вечером. Они либо планируют сюда из соседних участков леса, либо же приходят пешком, что особенно характерно в ветреную, ненастную погоду. В условиях Ленинградской обл. в разгар токования глухари слетаются на ток сразу же после захода солнца (Пукинский, Роо, 1966),но в пасмурную погоцу они могут появиться раньше. Впрочем, и в солнечную погоду появление глухарей на токовище может происходить в разное время.
Об этом же пишет и Вурм (Wurm, 1885), который сообщает, что глухари могут появляться на. токовище и до захода солнца, и при звездном небе. В тех местах, где глухарей не тревожат, им нет нужды удаляться далеко от токовища и они могут проводить весь день вблизи последнего или даже на нем (Крутовская, 1958; мои наблюдения в Ленинградской обл.). Многие принимают за «прилет» птиц на токовище начало демонстрационных полетов самцов (см. далее), так называемых подлетов. В действительности, птицы нередко прибывают на токовище несколько раньше.
Ход вечернего токования имеет существенные различия у разных популяций, и нам известно по крайней мере два типа вечернего поведения самцов. В одном случае (южная часть Карельской АССР, республики Прибалтики, Белорусская ССР и далее на восток южнее 60° с.ш.) вечернее токование начинается с характерных демонстрационных полетов. Самцы, собравшиеся на токовище, перемещаются, с дерева на дерево особым замедленным полетом, производящим громкий характерный шум, легко отличающийся от того, который производится нормально летящей птицей. Самцы определенно реагируют на звук такого полета, и стоит перелететь одной птице, как то же начинают делать остальные участники тока. Усаживаясь на ветку после такого «подлета», глухарь обычно издает гортанный хрюкающий звук, состоящий из трех слогов (вроде «хрр. . . хрр-хрр» или «у. . . э-у»). Птицы, летящие на токовище с кормежек, также заканчивают полет замедлением его с активной и шумной работой крыльев и характерным «хрюканьем» после посадки. Ближе к сумеркам, а иногда и сразу после «подлетов», количество которых у каждого самца колеблется от 2 до 5, самцы начинают петь. Сначала глухарь делает несколько начальных щелчков, оглядывается, снова начинает щелкать и так несколько раз. Спев первую песню, он начинает петь уже почти без перерывов, только временами делая небольшие паузы, чтобы оглядеться и прислушаться. Интенсивное пение продолжается до полной темноты, а на крайнем севере самцы могут петь в течение всей ночи до утра (Теплов, 1947а; Семенов-Тян-Шанский, 1959).
Собственно на этом вечерний ток заканчивается. Птица поет на избранном дереве, пока не засыпает. Во время вечернего пения самец может расхаживать по ветке, временами съедая пучок-другой хвои, или вдруг бьет клювом по ветви, производя глухие удары (мне этого наблюдать не удалось).
Несколько иной характер носит вечернее токование на севере европейской тайги (север Карельской АССР, Печоро-Илычский заповедник) и в Альпах. Здесь глухари, появляясь на токовище вечером, делают 1 —2 демонстрационных полета, а иногда обходятся и без них, после чего спускаются на землю или на снег. Они расхаживают по своим токовым участкам, поют, выполняют токовые взлеты и даже вступают в схватки друг с другом. Как отмечал Теплов (1947а), вечерние драки птиц на Северном Урале гораздо более частое явление, чем утренние, причем на шум, производимый дерущимися птицами, бросались другие самцы. Еще одна существенная деталь — появление самок на токовище с вечера. Они прилетают сюда примерно в то же время, что и самцы, а иногда и ранее и устраиваются ночевать на деревьях, не спускаясь на землю (Теплов, 1947а; Потапов, 1975). Возможно, что вечерним прилетом самок и объясняется повышенная активность самцов, хотя в Карельской АССР вечерних драк мне наблюдать не удалось. Очень показательно, что короткую ночь (полтора-два часа относительно темного времени) самцы спят на снегу, а не на деревьях (Потапов, 1975).
Утреннее токование проходит у всех .популяций примерно по одному плану. Оно начинается в южной половине ареала до рассвета (на 60° с.ш. примерна в 3 ч ночи). Самцы, просыпаясь, начинают щелкать и затем запевают. Необходимо заметить, что в отдельных случаях самцы могут появляться на току и в предутренних сумерках, особенно годовалые птицы. Интенсивность пения в это предрассветное время максимальная. Птицы поют почти непрерывно и исполняют за утреннюю зорю 600—700, а иногда и до 1300 песен (Пукинский, Роо, 1966; Muller, 1974). С началом рассвета, еще в густых сумерках, при освещении 3—5 лк прилетают или становятся деятельными ночевавшие здесь самки. Сначала они перелетают с места на место с характерной позывной (горловое, со слегка гнусавым оттенком «ак. ак. ак») и, если дело происходит в кульминационное время тока, приземляются на участках самцов, избранных ими в прошлые посещения.
С появлением самок самцы начинают один за другим слетать с деревьев вниз на землю. Различные сообщения о том, что в том или ином месте глухари предпочитают токовать на земле или, наоборот, на деревьях, есть результат сравнения различных периодов одного и того же токового процесса. В кульминационный момент самцы обязательно токуют на земле. В это время они особенно ревностно охраняют свои участки и немедленно устремляются на появившегося в их пределах соперника. Как правило, соперник в таких случаях убегает, не принимая боя, но иногда, особенно при стычках на границах соседних участков, схватки могут происходить довольно часто. В одних случаях драки происходят регулярно и носят упорный характер (Wurm, 1885; Hjorth, 1970; мои наблюдения в Ленинградской обл. и в северной Карелии). В других случаях, по многолетним наблюдениям, конфронтации между самцами-соседями никогда не доходили до драки (Muller, 1974).
Самки распределяются на току крайне неравномерно. У одного самца может находиться одновременно 4—7 самок, у другого же — ни одной. Нередко можно наблюдать, как целый табунок самок мирно кормится вокруг одного из самцов, который расхаживает вокруг них в основной токовой позе и исполняет песню за песней. С восходом солнца самки начинают разлетаться, часть самцов покидает участки вслед за ними, но наиболее активные продолжают токовать еще некоторое время на земле, а затем на деревьях. На не тронутых человеком токовищах в разгар токования самцы могут петь до 9—10 ч утра, причем некоторые из них остаются на земле и с окончанием токования начинают кормиться тут же, постепенно уходя пешком за пределы токовища.
Вопрос об индивидуальных участках самцов на токовище очень сложен и до конца не исследован. Данные многочисленных наблюдений противоречивы и свидетельствуют о большой индивидуальной изменчивости поведения самцов, а также об изменении степени территориальности одной и той же птицы в различные периоды токования. Наслаиваясь друг на друга, эти два фактора создают исключительное разнообразие в территориальном поведении глухарей.
В начальном периоде связь самца со своим токовым участком минимальна и четко не определена. Она возрастает с усилением токовой активности птиц и достаточно ясно, (учитывая индивидуальные отклонения в ту и другую сторону) проявляется в течение всего основного периода, когда самки посещают токовище. Особенно тесной эта связь становится в период спаривания и бывает максимальной вечером и на рассвете.
Индивидуальные вариации территориального поведения самцов проявляются как в величине токовых участков, так и в степени приверженности к ним. В отдельных случаях самцы могут менять токовые участки и токовища в течение одного весеннего сезона. Даже в течение одного утра можно наблюдать самые различные изменения территориальности птиц. На одном из токовищ Ленинградской обл. я наблюдал, как ток, начавшись на своем традиционном месте, после отлета самок внезапно переместился на две сотни метров в сторону, из болотистого сосняка на более сухое и открытое место, где самцы продолжали токование еще целый час. В другом случае, в северной Карелии 8 мая был найден ток, территория которого в последующие дни ежедневно перемещалась в одном направлении, так что за 6 дней ток переместился на 400 м (Потапов, 1975). В том же районе самцы на стабильных токовищах меняли свои участки при изменении условий: когда в результате бурного таяния снегов некоторые участки на болоте оказались затопленными, владельцы их переместились на соседние освободившиеся из-под снега склоны моренной гряды.
По этим причинам определить реальную величину токового участка довольно сложно, и, видимо, правы те авторы, которые считают, что у самцов нет четко фиксированных границ токовых участков и правильнее говорить о пограничных зонах, положение которых не отличается особенной стабильностью (Pirkola, Koivisto, 1970). На нейтральные зони, разделяющие участки, агрессивность самцов не распространяется, и именно по этим своеобразным каналам годовалые самцы могут проникать к самому центру тока. Взаимоотношения соседних самцов да и вообще вся структура иерархии глухариного тока, в особенности крупного, исследованы далеко недостаточно. Можно считать установленным, что в нормальных условиях центральные, ключевые участки на токовище занимают старые самцы, в возрасте от трех лет и старше.

Участки периферийные, менее удобные, занимают самцы в возрасте двух лет, а годовалые птицы держатся большей частью по краям токовища и своих участков не имеют.
Тот факт, что при наличии нескольких территориальных самцов самки концентрируются вокруг одного-двух из них, подчеркивает наличие такой трехступенчатой иерархии. Есть даже сообщения о наличии на току пятиступенчатой иерархии: 1-я ступень — самцы в возрасте менее 1 года (первогодки), 2-я ступень — двухлетки, 3-я ступень — трехлетки, 4-я ступень — четырехлетки (эти самцы названы «претендентами») и, наконец, 5-я ступень — доминанты, или «лидеры», которыми являются самцы в возрасте 5 лет и старше (Дронсейко, Немнонов, 1981). «Лидеры», занимающие самый верх иерархической лестницы, отличаются наиболее высокой активностью, что проявляется и в количестве песен — до 1300 песен за утро (Muller, 1974). Годовалые самцы, даже попадая в центр токовища, не имеют никаких шансов на успех. При наличии взрослых самцов самки полностью игнорируют молодых, хотя последние и пытаются.жреследовать самок. Примечательно, что и старые самцы почти не обращают внимания на молодых, что вызвано слабым развитием у последних вторично-половых признаков. Так, хвост молодого самца короткий, с узкими рулевыми перьями, будучи полностью раскрыт, не может образовать сплошной черной поверхности с контрастными белыми пятнами, как у старых птиц, а напоминает скорее половину колеса с торчащими спицами (рис. 100).
Небольшой темноокрашенный клюв молодого самца в отличие от клюва старой птицы совершенно не заметен в сумерках и не выполняет явно необходимой роли специфичного раздражителя (Потапов, 1975). Тем не менее, как уже говорилось, молодые самцы вполне половозрелы и в некоторых случаях, в основном при регулярном истреблении старых птиц, могут вступать в размножение. В таких ситуациях они занимают ключевые участки на токовище, активно поют и самки за неимением выбора вынуждены с ними спариваться.
Взаимоотношения взрослых самцов бывают подчас самыми необычными. Например, в вольерах Дарвинского заповедника наблюдали, как два самца дружно и слаженно токовали на одном участке, соблюдая полную синхронность как песен, так и всех движений — вместе начинали и заканчивали песню, одновременно двигались и поворачивались в одну и ту же сторону (Немцев и др., 1973). Поскольку аналогичное явление наблюдалось мною в природе у кавказского тетерева (см. далее), можно предположить, что такое явление имеет место у глухаря и в естественных условиях.
Согласно некоторым наблюдениям, началу спаривания предшествует довольно сложная предварительная фаза, в ходе которой территориальная структура тока полностью нарушается. Это происходит в то время, когда самки начинают приземляться в центральных участках токовища и расхаживают по нему небольшими группами или поодиночке. В такой ситуации сюда устремляются все находящиеся вокруг самцы и следуют за ними, нередко бок о бок, в основной токовой позе, образуя так называемый в охотничьей практике хоровод (Hainard, Meylan, 1935; Hjorth, 1970; Щербаков, 1967). Причем самцы не просто следуют за самками, но как бы стараются загнать их на свою территорию, и отдельным самцам временами удается привлечь на свой токовой участок сразу несколько самок. Во время этих действий границы участков самцами не соблюдаются и схватки между ними редки. В дальнейшем такого «хоровода» уже не происходит и самки приземляются уже на определенных участках отдельных, выбранных ими самцов. Окончательный выбор принадлежит только самке, и избирательность их в нормальных условиях очень высока. Об этом же говорят й наблюдения в вольерах (Немцев и др., 1973). Очень важно, что само спаривание, венчающее весь сложный токовой процесс, происходит именно на токовом участке самца и только тогда, когда имеется синхронная готовность к этому и самца, и самки. Спариванию обычно предшествует особая типичная для куриных птиц «приглашающая поза», принимаемая самкой. Есть сведения, что самцы непосредственно перед спариванием никогда не поют (Miiller, 1974), но по наблюдениям в вольерах, при щелканьи самца самка начинает подергивать крыльями, прижимается к земле и при «точении» принимает приглашающую позу (Кутовая, 1976). Спаривание следует вслед за этим далеко не всегда, поскольку самец в это время может быть отвлечен изгнанием соперника или же обхаживанием другой самки. По наблюдениям в Ленинградской обл., те самцы, вокруг которых собирается по 2—3 самки, спариваются каждое утро только с одной из них (Пукинский, Роо, 1966).
О том же сообщают Немцев с соавторами (1973). С.П. Кирпичев (устное сообщ.) полагает, что каждая самка спаривается за весну только один раз и этого бывает достаточно для снесения полной кладки. Однако в вольерах Березинского заповедника наблюдались повторные спаривания тех же партнеров через 20—30 мин и в течение периода спаривания каждая самка спаривалась неоднократно (Кутовая, 1976).
Заметные отличия в токовании глухарей были обнаружены в Западной Европе (Lumsden, 1961b; Hjorth, 1970), где численность птиц повсеместно низка, а наиболее подробные 15-летние наблюдения были проведены в одном из районов ФРГ, где этот вид был на грани исчезновения (Miiller, 1974). Все находившиеся здесь самцы по нескольку лет подряд занимали свои участки на токах и, кроме того, проявляли очень большую привязанность к территории в течение всего года. Вообще территориализм у самцов в деградирующих популяциях Западной Европы выражен гораздо сильнее, чем на остальной части ареала, вплоть до того, что самец охраняет участок с гнездящейся на нем самкой, т.е. ведет себя как моногам (Bannerman, 1963).
Следует особо подчеркнуть, что несмотря на нечеткую выраженность токовых участков, они тем не менее являются совершенно необходимым элементом в сложном механизме токового процесса как место, где только и может произойти спаривание. Некоторые исследователи, наблюдавшие как вслед за самками, покидавшими токовище, следовали иногда как молодые, так и взрослые самцы, предполагали, что спаривание может происходить вне тока. Однако еще никто такого не наблюдал, и если можно предположить возможность таких случаев, то станет совершенно непонятным существование столь сложного процесса токования с трехступенчатой иерархией. При нарушении естественных условий в вольерах при совместном содержании группы самцов, когда токует только «лидер»,.нетокующие самцы, в том числе и молодые, спариваются с самками даже чаще, чем токующие, но, возможно, с этим связано и заметное повышение процента неоплодотворенных яиц (Кутовая, 1976). Эти данные еще раз подчеркивают огромную генетическую роль сложного механизма тока глухаря, когда к размножению получают доступ только взрослые самцы.
Как только самки перестают посещать ток и приступают к насиживанию, активность самцов резко падает. Они еще продолжают появляться на токовище в течение 3—4 недель, причем прилетают сюда задолго до заката и подолгу кормятся на тех же деревьях, где и ночуют. После исчезновения самок они перестают спускаться по утрам на землю, и на зорях, как утренних, так и вечерних, поют все с меньшей активностью, перестают принимать во время пения токовые позы и вообще могут петь, не поднимая и не раскрывая хвоста. Как на севере, так и на юге ареала окончание пения глухаря фенологически совпадает с массовым ростом и раскрытием березовых листьев. Но в холодные затяжные весны песни глухарей можно слышать до начала—середины июня (северная Карелия, Урал, Красноярский край), когда у самцов уже начинается линька. Возможно, что самки, потерявшие кладку, могут вновь навещать ток для повторного спаривания, но конкретных данных на этот счет нет.
Для глухарей известно также летнее, осеннее и зимнее токование, наиболее выраженное на южных и западных окраинах ареала: По данным Кирикова (1952), на Южном Урале летнее токование происходит в первой половине августа. На места весенних токовищ собираются группы до 3 птиц, хорошо поющих и принимающих различные токовые позы. Все добытые здесь птицы были первогодками, линявшими во второй дефинитивный наряд, с увеличенными семенниками, где проходил активный сперматогенез, хотя последний и был выражен слабее, чем у взрослых самцов весной. Это наблюдалось также и в бассейне Рейна (Miiller, 1974), и в вольерах Березинского заповедника, где молодые птицы токовали уже в первой декаде июля. Взрослые самцы, первыми прекратившие токование весной, раньше завершали процесс активной линьки и вновь начинали токовать со второй половины августа (Кутовая, 1976).
Осеннее токование происходит в сентябре—октябре. Птицы поют как на деревьях, так и на земле в разное время на токовищах и на местах кормежек, часто в смешанных стаях, состоящих из самцов и самок. Один из самцов, добытых на таком току на Южном Урале, оказался двухлетним (Кириков, 1952). В бассейне Рейна самцы токуют на своих участках вплоть до конца ноября, причем их в это время регулярно навещают самки (Miiller, 1974).
Зимнее токование происходит значительно, реже осеннего и носит еще более случайный характер. В вольерах Березинского заповедника молодые глухари токуют всю зиму (Кутовая, 1976). Все эти явления правильнее рассматривать как проявление отдельных элементов токового поведения самцов (песня, черчение крыльями) в течение всего осенне-зимнего периода, с наименьшей активностью в середине зимы и ее постепенным усилением к весне. Так по крайней мере обстоит дело на Южном Урале и в Западной Европе (Wiirm, 1885; Zedlitz, 1929; Кириков, 1952, и др.). Состояние гонад в это время (сентябрь—ноябрь) в указанных местах не изучалось. Изучение состояния семенников у птиц с Кольского п-ова и северного Приуралья показывает отчетливое увеличение их в сентябре, хотя данные об осеннем токовании в этих местах очень скудны (Семенов-Тян-Шанский, 1959).
Брачный ритуал самцов довольно сложен. В основной токовой позе шея и хвост подняты вверх почти вертикально. Голова занимает более горизонтальную позицию, будучи поднята под углом около 45°. Перья на шее не топорщатся, и только удлиненные перья подбородка, так называемая борода, обычно стоят торчком.
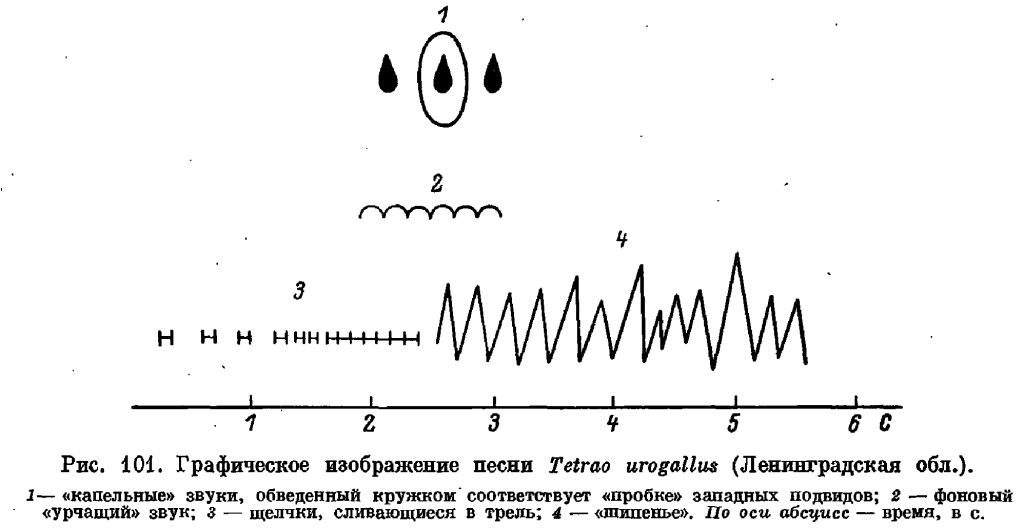
Концы крыльев приспущены, но не оставлены от тела, хотя в отдельные моменты они могут опускаться еще больше, а первостепенные маховые немного отставляться. В таком положении концы маховых перьев касаются грунта и при ходьбе волочатся по нему. Хвост максимально раскрыт, так что широкие опахала рулевых перьев образуют широкий полукруг (рис. 100).
Нижние кроющие перья хвоста также подняты, но под более острым углом, формируя специфический узор подхвостья. Во время наиболее активного периода токования, в густых сумерках, когда происходит спаривание, наиболее характерными для самца в основной токовой позе являются: а) характерный силуэт птицы с поднятыми шеей и хвостом и б) светлые (почти белые) пятна, контрастно выделяющиеся на сплошь темной в сумерках окраске самца — это прежде всего полукруг белых пятен на черном фоне раскрытого хвоста, белое «плечевое пятно» и светлое пятно массивного клюва. Как уже говорилось, степень развития белых пятен на рулевых перьях подвержена значительной индивидуальной изменчивости.
Песне глухаря посвящена весьма обширная литература (Wurm, 1885; Krüdener, 1928; Fuschlberger, 1956; Boback, 1966; Marchlewski, 1962; Hjorth, 1970, и др.), но до сих пор исчерпывающего ее описания составить нельзя из-за отдельных неясных моментов ее построения. Наиболее полное описание песни дает Хьорт (Hjorth, 1970), подвергший анализу синхронную запись движений и звуков, но и он не дает объяснения некоторым феноменам глухариной песни.
Сама песня глухаря — сравнительно тихая вокализация, слышная не далее чем на 150—200 м и длящаяся около 5.5 с (рис. 101). Она состоит из двух основных частей, резко различимых по звучанию и исполнению, причем первая часть в свою очередь может быть подразделена на две. Первая часть состоит из громких щелчков (или «тэканья»), сначала двойных, которые следуют друг за другом со все уменьшающимися интервалами от 0.5 до 0.13 с (первая часть песни), пока не сливаются в сплошную трель, длящуюся 0.5—0.8 с (вторая часть).
Трель эта внезапно обрывается короткой (0.1—0.2 с) паузой, вслед за которой следует фраза из звонких шипящих звуков, так называемое точение (из-за сходства со звуками, получаемыми при затачивании косы). Эта часть песни, длится около 3 с. Частоты песни лежат в пределах 1.5—6.5 кГц, с явно полифоническим звучанием в третьей части песни (выделяются полосы звучания на уровнях 2, 2.5 и 6 кГц, либо на 3.5 и 6 кГц).
В зависимости от степени возбуждения птицы начальная часть песни, состоящая из щелчков, может иметь разную длительность. Когда самец щелкает, как бы набирая разгон, и вдруг останавливается, затем после паузы начинает песню вновь и в конце концов запевает, то начальная часть содержит 6—8 двойных щелчков. Если же птица распелась и поет почти без перерыва, эта часть заметно сокращается, до двух-трех двойных щелчков, за которыми сразу же следует трель.
Там, где вторая часть песни («трель») переходит в «точение», вторым фоном прослеживается особый звук, накладывающийся на конец «трели» и начало «точения», слышный на небольших расстояниях (до 15 м) и, видимо, поэтому не отображенный на сонограммах. Этот глухой звук, похожий на урчание, очень удачно сравнивается со стуком гороха, сыплющегося в деревянное корыто (Lumsden, 1961b; Пукинский, Poo, 1966), И, наконец, на это урчание может в свою очередь наслаиваться еще несколько звуков, очень напоминающих звук падающей капли (рис. 101). У одних самцов, которых я слышал с небольшого расстояния в Ленинградской обл. и Карельской АССР, такой капельный звук был только один, у других их было три, а у некоторых он вообще не прослушивался. У южносибирских глухарей описано 2—5 таких звуков во время «точения» (Russow, 1924), а у шведских — 5 двойных, вроде «плип-плоп» (Hjorth, 1970). Обращает внимание явное сходство этих звуков с так называемым пробочным звуком («пробкой»), особым элементов, присущим песне самцов только трех западных подвидов — Т.u. urogallus, Т.u. major и Т.u. aquitannicus (Потапов, 1971). Название этого звука («пробка», «cork-note» англ., «соuрde-bouchon» — франц.) хорошо отображает его полное сходство со звуком при откупоривании бутылки (немецкие авторы называют его «Hauptschlag» — «главный удар»). Звук этот следует немедленно за «трелью», там, где у остальных подвидов имеется короткая пауза, и является самым громким в песне, слышным за 200—300 м. По своему звучанию он резко отличается от щелчков, слагающих две первые части песни. «Пробка» является новым элементом в песне глухаря, причем его развитие вполне могло произойти из первого «капельного» звука, произносимого как раз вслед за окончанием «трели». Действительно, на некоторых сонограммах этот звук выглядит как зачаточная «пробка». О молодости этого элемента в песне говорит его наличие у наиболее измененных западных подвидов глухарей (Потапов, 1971) и постепенность его развития в онтогенезе. По наблюдениям в неволё, самцы подвида Т.u. major поют в первую весну без «пробки» или же она очень слаба. Впоследствии год от года она становится все более громкой, и, по свидетельствам охотников, наиболее громкой и красивой «пробкой» отличаются самые старые самцы. При подходе к току западноевропейских глухарей первыми слышатся именно эти пробочные звуки токующих самцов, которые на большом расстоянии походят на звуки падающих в воду капель. Сходство этих звуков настолько велико, что было даже замечено определенное стремление у молодых глухарей токовать в первую весну там, где постоянно звучат капли талых вод, падающие со скал в лужи (Klaus et al., 1968).
Развитие песни у глухарей в процессе онтогенеза специально не исследовалось. При разведении в вольерах самцы проявляют способность к пению уже в первую осень, в возрасте 3—4 мес (первые попытки токования в возрасте 67—70 дней), хотя нормальный характер песня принимает в возрасте 6 мес. (Крутовская, Крутовская, 1953; Кутовая, 1976). Примечательно, что и у взрослых глухарей песня при токовании может значительно видоизменяться. Она может, особенно в разгар токования, почти полностью лишаться первой части (почти сразу начинается «трель»), либо же вместо третьей части самец заканчивает песню трехсложным хрюкающим звуком, либо этот хрюкающий звук заменяет первые две части. Чаще всего песня нарушается при токовых взлетах, следующих сразу за второй частью песни, — «точение» в таких случаях опускается. Эти аномалии указывают на возникновение глухариной песни из двух независимых компонентов — щелчков и «точения».
Последовательность движения челюстей при пении, характер движений в области горла и зоба, зафиксированные киносъемкой, не в состоянии объяснить происхождение тех или иных звуков песни. Несомненно, что в продуцировании звуков участвуют и сиринкс, и петля трахеи и язык птицы. Зоб не участвует в этом даже как резонатор, поскольку, по моим наблюдениям, птица может петь с зобом, наполовину заполненным хвоей. Важно подчеркнуть, что щелканье производится не клювом (заблуждение, которое до сих пор встречается в литературе), а сиринксом. При щелчках клюв вообще ни разу не смыкается полностью. При первом звуке двойного щелканья нижняя челюсть слегка приоткрытого клюва резко опускйетСя вниз и медленно поднимается обратно со второй нотой. Одновременно с этим происходит выдох. Спустя 0.1 с после движения челюсти вниз горло слегка втягивается, сразу же возвращаясь в начальную позицию, и вторично совершает это движение спустя еще 0.3 с после второй ноты. В процессе двойного щелканья совершают движение и язык с ларинксом и верхней частью трахеи, втягиваясь внутрь примерно на 5 см и возвращаясь назад, что хорошо видно по вздрагиванию вентральной поверхности верхней чаісти шеи. Столь же отчетливо его левая часть груди как бы втягивается с первой нотой и возвращается в начальную позицию со второй (на зобе слева, как было показано выше, покоится петля трахеи). При учащении «теканья» и при трели голова самца поднимается во все более вертикальную позицию и в момент «пробки» даже запрокидывается назад. Сам «пробочный» звук произносится в момент, предшествующий смыканию челюстей, — они смыкаются в первый раз во время пения при откидывании головы назад.
Это происходит в доли секунды, после чего голова возвращается в свою первоначальную позицию (одновременно с открыванием клюва) и начинается заключительная часть песни — «точение». Самец начинает его со слегка открытым клювом и в течение 3 с, пока оно длится, полностью закрывает его 7 раз. Во время «точения» голова и шея судорожно подергиваются то вперед, то назад. С каждым открыванием клюва из него вырывается воздух (что хорошо видно при морозе), а вентральная поверхность шеи в центральной ее части дважды слегка вздувается на третьей секунде звучания. В исполнении «точения» активное участие принимает и язык. Общеизвестен тот факт, что у добытых в эти моменты самцов язык оказывается глубоко втянутым в глотку. Кроме того, вибрация горла во время «точения» указывает на быстрые возвратные движения языка именно в этой области.
Самцы подвидов, у которых в пении нет элемента «пробки», при переходе к третьей части песни не откидывают голову назад и клюв не достигает вертикальной позиций. Предположение, что своеобразное урчание, накладывающееся на конец «трели» и начало «точения», продуцируется быстро повторяющимся смыканием и размыканием челюстей (Пукинский, Роо, 1966), не подтверждается анализом кинокадров. Скорее всего этот звук происходит при перемещениях воздуха в сообщающейся системе трахея—эзофагус. В отношении же «пробки» можно только предполагать, что здесь происходит процесс, аналогичный тому, что имеет место при откупоривании бутылки, т.е. происходит заполнение вакуума. Но, где образуется этот вакуум, пока неясно, а роль «пробки» выполняет, видимо язык.
Явление «глухоты» глухаря при «точении» широко известно и с незапамятных времен используется охотниками для приближения к поющей птице на близкое расстояние. Глухарь перестает реагировать на звуки в последнюю секунду второй части песни («трели») и в течение почти всей третьей части, однако же в самые последние моменты «точения» он вновь обретает слух. Таким образом, невозможно связывать «глухоту» глухаря только с одним коленом песни. Вообще, видимо, следует говорить не о полной, а только о частичной утере слуха. Опыты с самцами, токовавшими в условиях неволи (Marchlewski, 1962), показали, что глухари все же слышат во время «точения». Так, они прерывали его при близких выстрелах, при звуке трубы, при барабанных ударах. Частичная же потеря слуха объяснялась тем, что расположенная в наружном ухе небольшая железа во время пения наливается кровью, расширяется и перекрывает слуховой проход (Wurm, 1885). Впоследствии предполагалось, что замыканию слухового прохода этой железой способствовало движение сочленового отростка нижней челюсти, к которому крепится пратнерова связка, идущая от слухового столбика (Ильичев, 1964). Проверить это можно только в опыте методикой вживленных электродов, что до сих пор сделано не было. Но основное сомнение в правильности этой гипотезы вызвано тем фактом, что в период «точения», когда птица глохнет, клюв самца вообще закрыт и челюсти не двигаются.
Во время пения самцы могут принимать самые различные позы, но обычно в разгар токования поющая птица распускает полностью хвост (рис. 100) и поднимает его почти вертикально. Крылья слегка распускаются и отводятся в стороны и вниз, шея и голова подняты вверх. Начиная пение в предутренних сумерках, самец поначалу может раскрывать хвост далеко не полностью, лишь во время «точения», и высоко его не поднимает, крылья только слегка приспущены. С рассветом позы поющего самца становятся все более напряженными, глухарь как бы входит в азарт, становится более подвижным. Собственно в это время он и принимает наиболее характерную для пения позу, описанную выше.
Шея при пении не раздувается, хотя вообще в весенний период она заметно расширена из-за существенного утолщения подконщой клетчатки, сильно набухающей и переполняющейся серозной жидкостью (Теплов, 1947а). Такое утолщение несомненно несет защитную функцию, предохраняя шею от сильных ударов клювом во время турнирных боев. На дереве поющая птица обычно стоит на одном месте или же переступает с ноги на ногу, поворачиваясь в разные стороны. Однако на земле глухарь может во время пения довольно быстро перемещаться и-даже взлетать, прерывая при этом песню. Во время сильного дождя или ветра, в конце периода токования, а также осенью самцы могут петь, не поднимая и не раскрывая хвоста.
На большей части ареала для глухарей характерны особые токовые полеты, так называемые подлеты, исполняемые вечером, перед началом вечернего пения. Появляясь на токовище, самец обнаруживает свое присутствие именно этим полетом, который совершается по прямой в замедленном темпе и сопровождается усиленно громким биением крыльев, нередко задевающих при этом за окружающие ветви. Такой полет, если он первый в данный вечер, может начинаться с земли, но заканчивается обязательно на дереве. Усаживаясь, птица первым делом издает гортанный «хрюкающий» звук (см. далее). Токовой полет обычно выполняется в пределах своего токового участка, и «территориальная» его функция очевидна. Обычно за вечер самец проделывает 2—3 таких полета, но число их может значительно увеличиваться на току с большим количеством птиц, где самцы провоцируют такими полетами друг друга. Расстояние токового полета небольшое, в среднем около 50 м, и совершается он на высоте в полдерева. Этот ритуал, позволяющий довольно точно определить вечером количество самцов, участвующих в токовании на данном токовище, широко используется охотниками как предварительная рекогносцировка перед утренней охотой (так называемый подслух). Утром, если самец ночевал на дереве, совершается обычно только один токовой полет — с дерева на землю, обычно в центр, токового участка или же к самкам, приземлившимся поблизости.
Токовой взлет совершается на земле, причем он характерен как для начала, так и для наиболее активной фазы тока. Токовой взлет одного самца стимулирует аналогичное поведение других. Взлет может стимулироваться также появлением самки или ее отлетом (Lumsden, 1961). Нередко взлет выполняется во время пения, заменяя последнюю часть песни. С места, или сделав несколько шагов, птица полого взлетает на высоту полутора-двух метров и столь же полого опускается с громким хлопаньем крыльев. Никаких голосовых звуков при этом не издается. При взлете делается 4—5 сильных коротких взмаха, затем наступает краткая пауза, во время которой глухарь как бы замирает на мгновение с распростертыми крыльями в верхней точке полета, и вслед за этим он опускается, делая вновь несколько взмахов. По моим наблюдениям в северной Карелии, число взмахов при взлете и спуске может быть различным — на спуске число взмахов заметно меньше. Со стороны токовые взлеты воспринимаются на слух как два взрыва хлопков (на взлете и посадке), разделенные паузой в доли секунды. По моим измерениям, глухарь пролетает во время таких взлетов от 3 до 7 м, обычно по прямой, но нередко в воздухе он меняет направление на 45—90°. Во время первого взмаха крылья бьют по земле или по снегу, оставляя на последнем отпечаток, но приземление идет более круто, без касания крыльями земли, сразу на обе ноги (толчок перед взлетом, если таковой и делается, выполняется вразножку). Иногда при приземлении в рыхлый снег самец по инерции падает на него грудью и на короткий миг замирает в такой позе с расправленными крыльями и полуоткрытым хвостом, что четко отпечатывается на снегу — совершенно так же, как у азиатской дикуши после токового прыжка (Потапов, 1969а). Впрочем, такое падение на грудь наблюдалось после посадки и на твердом субстрате.
Кроме пения самцам свойственна еще одна особая вокализация — «скирканье», — которое издается на утреннем и вечернем току и обычно характерно для заключительного периода токования. Оно состоит из щелчка, за которым следует особый скрежещущий звук, как при чирканьи косы о камень, вслед за чем следует еще один щелчок. Поза глухаря при «скирканьи» весьма характерна — хвост раскрыт и поднят вертикально, голова и слегка изогнутая шея вытянуты вперед и во время «скирканья» совершают короткий поклон, маховые слегка отставлены. «Скирканье» наблюдается нечасто, оно может издаваться самцами и на лету, а осенью они также могут «скиркать», будучи подняты Собакой, «Распластывание» — особая поза, принимаемая самцами по крайней мере в первый и второй периоды токования, была известна только по отпечаткам на снегу. Самец, расхаядавая по снегу, «. . .иногда прижимает к его поверхности вытянутую шею, распущенные крылья и хвост. Если наст покрыт свежей порошей, то на снегу в таких случаях остается чрезвычайно своеобразный отпечаток контуров всей птицы» (Теплов, 1947а). Такие отпечатки я встречал в Ленинградской обл. довольно рано, уже с середины февраля вместе со следами токовых взлетов. Большей частью на снегу остается отпечаток расправленных крыльев, наполовину раскрытого хвоста и туловища до основания шеи включительно. Саму позу мне удалось впервые наблюдать в северной Карелии. Один из самцов, расхаживая на своем токовом участке, изредка ложился на снег грудью, вытянув вперед и вверх голову. Крылья его были сложены, а хвост наполовину раскрыт и опущен, но один раз самец проделал этот ритуал, распластав крылья по снегу. В такой позе птица находилась несколько секунд.
Очень редким элементом брачного ритуала является токовой прыжок. Описание его имеется только у Теплова (1947а). Птица подпрыгивает на высоту около 30 см, поджимая крылья, закидывая назад голову и издавая каждый раз хриплый отрывистый звук. Прыжки наблюдались как по утрам в начальный период токования, так и в разгар его по вечерам.
Схватки самцов бывают очень ожесточенными. Им предшествуют своеобразные позы угрозы. Начав кружить друг против друга, соперники делают угрожающие выпады клювом и временами бегут друг другу навстречу. Хвост при этом приподнят и сложен, голова и шея при беге навстречу противнику быстро двигаются вверх и вниз. Оказавшись друг перед другом, самцы принимают дозу, несколько отличную от основной: хвост распущен и Поднят под углом 70—90°, шея стоит вертикально и слегка втянута, но голова держится почти параллельно земле. Пррья «бороды» стоят торчком, а перья шеи и верхней части груди взъерошены, благодаря чему шея кажется сильно расширенной (рис. 100) и более темноокрашенной, так как серые кончики перьев стоят вертикально, открывая темную окраску оснований. Из этой позиции птица, издав гортанное «хрюканье», делает быстрый поклон, так что голова почти касается земли, и в такой позе повторяет «хрюканье» более быстро. Этот маневр занимает около 2 с, и стоящие друг против друга самцы могут выполнять его синхронно. Конфронтирующие самцы нередко тоже синхронно делают быстрые поклоны и угрожают друг другу, то вытягивая шею по направлению к сопернику, то втягивая ее обратно.
Иногда конфронтирующие самцы могут принимать особую позу, ложась грудью на землю и слегка распластывая крылья. Хвост в такой позе по-прежнему раскрыт и поднят вверх. При близких конфронтациях, обычно предшествующих схватке, соперники то и дело издают гортанный хрюкающий звук из 4 слогов, длящийся 0.9 с, причем первые два слога звучат почти слитно. Диапазон этих звуков лежит в пределах 2—7 кГц, и хотя их сонограмма очень сходна с сонограммой песни, звучат 1они совсем иначе и походят на глухое хрюканье. Обращает также на себя внимание сходство ритмики этого «хрюканья» с криком фазана — те же два двойных звука, где два первых почти сливаются.
Нападающий самец изгибает шею дугой, складывает и слегка поднимает хвост и частично взъерошивает перья на шее (рис. 100). При этом самец издает своеобразный шипящий звук «фффффф», иногда заканчивающийся обычным «хрюканьем» (Потапов, 1975). Во время драки птицы хватают друг друга клювами за шею, бьют крыльями. Южноуральские глухари, сходясь, поют непрерывно, затем замолкают и кидаются в драку (Кириков, 1952). Схватки бывают жестокими, птицы получают серьезные ранения, шрамы от которых, особенно на шее, сохраняются довольно долго, а по данным западных авторов (Boback, 1966; Hjorth, 1970), некоторые птицы получают столь тяжелые раны, что погибают на месте драки, хотя такие случаи и редки. Дерущиеся глухари теряют всякую осторожность, они могут подпускать человека вплотную и даже позволяют схватить себя руками. Бой заканчивается, как правило, победой хозяина участка и бегством побежденного, которого победитель преследует на некоторое расстояние. Во время вечерних драк на шум крыльев сцепившихся самцов кидаются другие глухари, но принимает ли при этом баталия коллективный характер, не сказано (Теплов, 1947а). Сражения глухарей носят несомненно турнирный характер,и ряд приспособлений как морфологического (утолщение шеи), так и этологического порядка (оставление соперником участка хозяина) предотвращает, как правило, серьезные увечья или даже смертельный исход для соперников.
Самка выбирает для гнезда место недалеко от токовища, причем иногда две самки могут гнездиться в непосредственной близости одна от другой в 40—50 м (Белоруссия; Долбик, 1961). Гнездо обычно прикрыто свисающей еловой ветвью, кучей хвороста, упавшим стволом, выворотом корня или же помещается под камнем. Нередко гнездо устраивается у основания ствола березы или сосны, обычно с южной стороны и гораздо реже просто в густой траве или посредине поляны или перелеска. Само гнездо — небольшое углубление в почве, слегка выстланное сухими листьями и травинками, отдельными перьями, обрывками мха. Почва под гнездом обычно сухая. Выстилка в гнезде постепенно накапливается в процессе откладки яиц. Самка собирает клювом перед собой хвоинки, ветошь, кусочки мха-и характерным движением забрасывает их себе за спину (Кутовая, 1976). При задержке весны самка может начать откладку яиц прямо на снег. Описан случай, когда самка отложила первое яйцо 19 июня (северная Финляндия 61°15′, с.ш.) на снег под кроной упавшей сосны. Средняя глубина снега в это время вокруг гнезда была около 50 см, хотя имелись уже и бесснежные пятна. Первые 4 яйца были снесены прямо в снег, и только после снесения четвертого яйца под них был подложен мох. К моменту откладки 9-го яйца снег вокруг растаял (Pulliainen, Rajala, 1973). Такие случаи, видимо, не составляют редкости в северных частях ареала, где в отдельные годы обилие снега или холодная весна сильно задерживают его таяние.
Окраска яиц значительно варьирует, даже у птиц из одной области, а кроне того, цвет яиц существенно меняется в процессе насиживания. Общий тон окраски палево-песочный или слегка коричневатый, по которому густо и равномерно разбросаны мелкие темно-коричневые пятнышки. Среди них обычно бывает несколько пятен покрупнее, диаметром до 2, реже до 3—4 мм. Встречаются очень светлые яйца, с почти белым фоном и светло-коричневым цветом мелких пятен, и, наоборот, густо пигментированные, где многочисленные коричневые пятна сливаются в почти сплошные поля. Цвет яиц явно зависит от характера питания самки: пигментация тем гуще, чем больше в пище молодых побегов и почек хвойных, и тем бледнее, чем больше в питании зерновых кормов (С.П. Кирпичев, устное сообщ.). Размеры яиц с Кольского п-ова (29): 57—6І х 41 —43 мм, средняя масса 53.2 г (Семенов-Тян-Шанский, 1959); из Печоро-Илычского заповедника 58.4 x 41.2 мм (56.2—60.5 x 40.5—42.1) (Теплов, 1947а). Размеры 22 яиц из коллекции ЗИН АН СССР —57.2×41.7 мм (54—63.7 х X 39.1—46). Среди них особенной величиной отличается яйцо глухаря из-под Барнаула — 63.7 x 46 мм. Несомненно, что величина яйца зависит и от возраста -самки. Толщина скорлупы в средней части яйца — 0.4 мм.
Количество яиц в кладке варьирует даже в одной и той же местности, находясь в зависимости и от возраста самок и от характера зимы и весны. По данным Кирикова (1952), на Южном Урале в благоприятную весну 1946 г. были очень крупные кладки, одна из найденных кладок насчитывала 16 яиц, а в выводках наблюдалось по 12 молодых. Там же в холодные дождливые весны кладки содержали примерно по 5 яиц. Возможно, что крупные кладки образуются при одновременной откладке яиц двумя самками в одно гнездо. В вольерах это во всяком случае наблюдалось (Кутовая, 1976).

Данные табл. 45 показывают в общем примерно равную плодовитость по всему ареалу, но в Финляндии происходит уменьшение величины кладки от 7.7 на юге до 5.8 на севере (Linden, 1983а).
В Швеции самка приступает к откладке яиц через 3 дня после спаривания л несет ежедневно по яйцу (Hoglund, 1952). По Семенову-Тян-Шанскому (1959), промежуток между откладкой яиц больше чем сутки (самка снесла 5 яиц за 6 сут).
По наблюдениям в вольерах, самки начинают кладку через 3—4 дня после спаривания. Самые короткие интервалы между откладкой яиц — 30—33 ч, более обычны интервалы в 46 —48 ч и очень редко, обычно в конце кладки, бывают интервалы в 4 сут (Кутовая, 1976). С откладкой каждого яйца птица сидит на гнезде все дольше и окончательно приступает к насиживанию перед снесением последнего яйца. Самка насиживает очень плотно, покидая гнездо в среднем дважды в сутки, утром и вечером, реже 3 раза и еще реже 1 раз в сутки. Отлучки длятся 10—35 мин, в теплую погоду дольше, вхолодную меньше. Самка может оставлять кладку и при легких заморозках, что никак не отражается на развитии эмбрионов, но она всячески избегает покидать гнездо во время дождя. При затяжных дождях самка сидит на гнезде непрерывно, но более полутора суток, видимо, выдержать не может. Самка регулярно переворачивает яйца, около 3 раз в сутки, а в ненастье 6—9 раз (Семенов-Тян-Шанский, 1959).
В Шведской Лапландии (Lennerstedt, 1966) в холод и снегопады самки оставляли гнездо чаще, 4—5 раз в сутки. Видимо, в холодную погоду самка вынуждена чаще кормиться, чтобы восполнить энергетические потери, возрастающие с падением температури. Отлучаясь, птицы не прикрывали кладку. В Финской Лапландии самка, находившаяся под наблюдением телекамеры, не прикрывала кладку при отлучках, если не считать нескольких сухих стебельков, бросаемых на яйца. Температура воздуха в период насиживания колебалась в пределах —0.5 и 29 °С. Здесь в условиях непрерывного дня также наблюдались 1—3 отлучки в сутки, чаще в вечернее время и никогда в интервале между 13 и 19 ч. Средняя продолжительность отлучек 34 мин (20—70 мин). Самка регулярно, с интервалами в среднем в 100 мин, передвигала яйца боком клюва. Птица покидала гнездо пешком и, пройдя 1 м, взлетала обычно в восточном направлении, заворачивая затем на север. Место кормежки располагалось в 500 м к северо-востоку. Здесь самка усиленно питалась листьями и бутонами черники и, наполнив зоб, возвращалась назад, приземляясь в 2—5 м от гнезда и подходя к нему пешком, предварительно осматривая местность. При вылуплении птенцов самка перемещала скорлупки внутри гнезда, вкладывала один кусок в другой, но не удаляла их из гнезда. Все птенцы вылупились в течение одного дня, и на следующий день рано утром покинули гнездо вместе с самкой на 16 мин, затем вернулись обратно в гнездо, где самка обогревала их около получаса, и окончательно покинули гнездо еще через 20 мин (Pulliainen, 1971). Но наблюдениям Крутовской и Крутовской, самки, насиживавшие кладки в вольерах, оставляя гнездо, закрывали кладку мхом или сухими листьями. По данным Кутовой (1976), при насиживании в вольерах далеко не все самки закрывали кладку при отлучках.
В Альпах самка ежедневно оставляла гнездо трижды: около 4 ч утра, между 11.30 и 15.00 и вечером в 19.00 или попозже, в сумерках, отсутствуя каждый раз в среднем 20 мин. Собираясь оставить кладку, птица делалась беспокойной, многократно меняла свое положение и, поднявшись и постояв с минуту, шагала в сторону и быстро слетала вниз по косогору. Только раз самка покинула гнездо пешком. Возвращаясь, самка останавливалась метрах в 10, быстро оглядывалась и потом уже бежала к кладке. Примерно дважды в день самка проделывала на гнезде процедуру купания, как на порхалище, только без хлопанья крыльями (Krott, 1966).
Самка насиживает очень плотно, иногда подпускает человека вплотную, не взлетая, а буквально отползая от гнезда (Кириков, 1952). Но так бывает не всегда, и большей частью при опасности самка взлетает «свечкой» с громким хлопаньем крыльями.
По данным Грюнера (1951), максимальные температуры насиживания колебались в пределах 34.8—43.6° С, а минимальные 14.6—23.7 °С. Яйца, оставленные ушедшей на кормежку самкой, при 17 °С остывали за полчаса только на 5°, а при температуре среды 0°С — всего на 10°. Периодические охлаждения, видимо, вообще необходимы для нормального развития зародышей. Данные различных авторов о режиме насиживания сведены в табл. 46.
Режим насиживания зависит и от возраста, и от индивидуальных особенностей самки, и от погодных условий во время насиживания.
Длительность насиживания определяется разными авторами в 20—30 дней, в среднем 26 дней (Hoglung, 1952; Семенов-Тян-Шанский, 1959; Ивантер, 1965; Pulliainen, 1971). Колебания эти явно зависят от режима насиживания. Срок в 20 дней (Ушков, 1887) кажется слишком коротким и другими данными не подтвержден.
Масса только что вылупившегося птенца 35.8 г (Карелия; Ивантер, 1965), 36—37 г (Кольский п-ов; Семенов-Тян-Шанский, 1959), 33 г (Кировская обл., Ефремов, 1940), 38—45 г (Красноярский край; Крутовская, Крутовская, 1958), 38.2 г (Швеция; Hoglund, 1955). Столь значительные колебания (от 33 до 45 г) говорят о явных различиях в массе яиц и несомненно сказываются на общем успехе репродукции популяции.

После вылупления самка обогревает обсыхающих птенцов, и в зависимости от погоды они либо сразу покидают гнездо, либо проводят там еще несколько часов. Первые 1—2 дня птенцы существуют за счет желточного мешка, хотя с первых же часов жизни активно ищут и поедают насекомых. Масса птенцов в это время не растет и может даже снижаться в первые сутки на 2—3 г (Hoglund, 1955; Семенов-Тян-Шанский, 1959), поскольку в это время они существуют главным образом за счет желточного мешка. Поиск пищи с первого дня полностью инстинктивен. Птенцы клюют движущихся насекомых, капли росы, яркие пятна, причем да насекомых они склевывают с нижней поверхности листьев.
В первые дни птенцов особенно привлекает зеленый цвет, а из предложенных растений они методом проб и ошибок быстро избирают цветки (Hoglund, 1955). Опыт разведения в вольерах показал, что тот корм, который будет предложен птенцу в первые 6 ч жизни, становится для него впоследствии наиболее предпочтительным. Но если в течение первых 24 ч птенец корма не получил, то в дальнейшем он поедает только мелких насекомых (Кутовая, 1976).
Полная зависимость благополучного развития выводка от погодных условий доказана экспериментально (Hoglund, 1955). Температура тела у только что вылупившихся птенцов 37.9° С. Она постепенно поднимается и достигает 41.6°С в возрасте 18 дней, когда птенцы приобретают способность к поддержанию постоянной температуры тела. Эта температура остается уже неизменной, испытывая лишь незначительные суточные колебания. В первые же дни жизни птенец теряет тепло очень быстро. На рис. 102 показана минимальная температура воздуха, при которой птенцы могут кормиться, не снижая активности, в течение 20 мин (в условиях сухой погоды). Из этого рисунка становится совершенно ясной необходимость постоянного подогревания птенцов самкой. Чем ниже температура воздуха, чем выше влажность, тем быстрее охлаждаются птенцы, тем чаще и дольше они должны проводить время под самкой, восстанавливая потери тепла, и тем меньше времени им остается на добычу корма, а количество насекомых в холодную погоду сильно уменьшается. Вполне очевидно, что при длительных холодах с дождями птенцы быстро погибают от истощения.
В тех редких случаях, когда, несмотря на ненастье, корм имеется в изобилии, птенцы благополучно переживают неблагоприятное время. Так, например, у берегов Байкала, несмотря на дождливую холодную погоду, птенцы успешно с первых же дней переживали ее, используя изобилующий корм — личинки хирономид (Кирпичев, 1968). Этот же автор отмечал, что птенцы в первые дни жизни не намокали под дождем благодаря наличию функционирующей копчиковой железы. В то же время Хёглунд утверждает, что в первые дни жизни копчиковая железа у птенцов не функционирует и, несмотря на то что какое-то количество секрета попадает на птенцов с оперения самки, они намокают под дождем довольно быстро.
Ненастная погода действует отрицательно и на птенцов более старшего возраста. Полуторамесячные птенцы при затяжных дождях могут вдвое снижать суточные привесы и замедлять линьку, даже несмотря на подкормку (Кирпичев, 1968).
Развитие птенцов происходит следующим образом. В возрасте 4—5 дней они ужё начинают перепархивать, на 5-й день они могут взлетать на высоту 30 см, а на 8-й день до 1 м (Крутовская, Крутовская, 1953; Krott, 1966). Регулярное «купание» в сухой трухе было отмечено с 7-го дня, поедание растительного корма с 11-го дня, реакция на хищников в воздухе — с 13-го дня, в возрасте 17 дней птенцы могли пролетать 10 м (Krott, 1966). В природе и при вольерном разведении у птенцов в первые 10 дней жизни очень силен инстинкт затаивания при опасности — птенец не двигается даже если его трогают (Теплов, 1947а; Крутовская, 1958). Затем на смену затаивания в возрасте 10—12 дней приходит реакция взлета. Взлетев, птенцы стараются улететь как можно дальше и снова садятся на землю, где затаиваются. С появлением ювенального наряда, т.е. с трехнедельного возраста, птенцы, как правило, взлетают на деревья. Оперившиеся птенцы, садясь на дерево, вытягивают шею горизонтально вдоль сучка и затаиваются, сливаясь окраской с веткой, особенно, когда они садятся на сосну. Затаившись, глухарята не слетают, несмотря на любой шум и выстрелы (Апраксин, 1900; Ефремов, 1940). Самка проявляет большую заботу о выводке вплоть до того времени, пока он окончательно не станет на крыло.
Первый месяц выводок проводит, как правило, в ельниках, по окраинам вырубок и особенно верховых болот, где более всего предпочитается полоса тростников вдоль невысоких грив, заросших лесом. В первый месяц жизни очень часто посещаются заливные луга, опушки и ноляны — места, наиболее богатые насекомыми. Распад выводка начинается примерно с 80-го дня жизни, с появлением агрессивности у самцов и их начальных попыток покрывать самок (Крутовская, Крутовская, 1953; Krott, 1966). По наблюдениям Кутовой (1976), иерархические отношения в выводках неустойчивы и могут легко меняться. Поза доминирования начинает появляться с 6-дневного возраста, а первые попытки токования отмечались у самцов в возрасте 67—70 дней. Остатки выводков, состоящие из молодых и старых самок, к зиме собираются в стаи, иногда большие. Самцы образуют отдельные группы, которые поздней осенью могут соединиться со стаями самок.
О размерах и массе растущих птенцов дают представление таблицы и графики, составленные Крутовской и Крутовской (1953), Тепловым (1947а) и Семеновым-Тян-Шанским (1959). Некоторый разнобой в данных объясняется приблизительным определением возраста птенцов на воле. Кроме того, в различных частях ареала скорость роста молодых птиц различна, будучи выше на севере. В конце первой осени молодые птицы, в особенности самцы, далеко еще не достигают размеров взрослых птиц.
Летняя жизнь самцов сравнительно однообразна. Они начинают линять уже в конце токового периода и в период интенсивной линьки (июль—август) стремятся обитать в самых глухих захламленных участках леса, обычно со значительной примесью ели, где много хвощей, папоротников и осоки. Ближе к осени они перебираются на ягодники (на морошку, чернику или землянику) и позднее, в августе, на брусничники. Большую часть этого времени глухари мало подвижны и держатся на земле, где иногда и ночуют, хотя чаще ночевки бывают на деревьях. Ушков (1887) весьма точно описал летние повадки линяющих самцов: « В это время летает тихо и поднимается вверх для полета с разбега, как журавль. От преследования собаки старается убегать в кустарник, мшарник, под вершину срубленного дерева и, забравшись туда, выдерживает мертвую стойку собаки. Вообще, в период линяния глухарь особенно не любит, когда его тревожат и ленив на подъем ». Как самцы, так и холостые самки держатся поодиночке, по двое, реже собираясь в небольшие группы одного пола, по 5—6 птиц. Это как бы основа осенней стаи, к которой впоследствии присоединяются птицы из других выводков. Годовалые птицы ведут себя так же, за исключением времени летних токов в южных районах, описанных выше. Мне в конце, лета приходилось встречать старых самцов на брусничниках, обычно по 1—2, птицы.
Для летнего времени очень характерны регулярные посещения птицами порхалищ, расположенных в определенных местах, обычно на солнцепеке. Это либо выходы песчаного грунта на тропе, на склоне или на месте выворота упавшего дерева, либо труха на месте сгнившего ствола, либо даже рассыпавшаяся завалина лесной избушки. Птицы посещают эти порхалища регулярно, независимо от пола и возраста, в солнечные и жаркие дни около полудня. В южных частях ареала, где лето бывает засушливым, как, например, на Южном Урале, птицы во время засухи концентрируются у воды, где и проводят жаркую часть дня.
Хозяйственное значение глухаря определяется как его биоценотической ролью в экосистемах бореальных лесов, так и тем важным местом, которое он занимает в охотничье-промысловой и спортивной охоте.
Благодаря значительным размерам самих птиц и периодически достигаемой, ими высокой численности глухари являются существенным компонентом бореальных лесных экосистем, роль которого особенно возрастает зимой. Как консумент он вводит в ускоренный кругооборот веществ значительное количество, растительной массы, преимущественно хвои, являясь основным ее потребителем, в зимнее время. Две трети этой массы, поедаемой птицами, возвращается ими обратно в виде экскрементов. При средней численности птиц, равной 52.3, на 1000 га (при равном соотношении полов), за 7 месяцев холодного сезона это, составляет 1624.3 кг экскрементов, а при максимальной численности вида — до 6497.4 кг (Потапов, 1981). Все это количество экскрементов консервируется, в снегу и одновременно поступает в почву весной, с началом вегетации растительности. Сколь ни казалась бы незначительной эта цифра, роль этих органических удобрений в условиях исключительной бедности органикой подзолистых почв северных лесов несомненно положительна. В местах же концентрации птиц (например, на токовищах) повышенное количество экскрементов, вызывает обогащение травянистой растительности (Казанцева, Морозкин, 1981). Потребляя большое количество хвои и концевых побегов ветвей (около 5 т на 1000 га), глухарь в то же время не наносит заметного урона лесу. И если в ряде, случаев кормежка птиц на избранных «кормовых» деревьях может причинять, последним повреждения и даже вызывать их гибель, то в общей древесной массе фитоценоза такие потери не превышают долей процента.
Биомасса глухаря при том же среднем уровне численности составляет 156.9 кг, и во многих случаях на долю одного этого вида приходится более половины биомассы всех птиц, зимующих в данной экосистеме. При максимальной же численности биомасса вида возрастает до 870 кг на 1000 га. Глухарь является поэтому немаловажным источником питания для целого ряда хищников, среди которых находятся и такие важные для пушного промысла виды, как куница, соболь, лисица.
Хозяйственное значение и охрана. Роль глухаря как объекта промысла никогда не была особенно высокой даже в.прежние времена, когда численность этого вида повсеместно была значительной. Это объясняется прежде всего гораздо более низким качеством мяса глухаря по сравнению с остальными тетеревиными птицами. Сейчас, правда, в силу резкого повышения спроса и цен на любую дичь этот вид также стал предметом экспорта, но теперь уже его промысловым заготовкам во многих областях препятствует низкая численность. Поэтому сейчас глухарь наиболее перспективен как объект спортивной охоты для всех без исключения охотничьих хозяйств таежной зоны. Его крайняя оседлость, прекрасная адаптированность к местным условиям, большой репродуктивный потенциал и высокий предел плотности населения делают этот вид исключительно выгодным для спортивного охотничьего хозяйства. Он уже давно эксплуатируется в ряде стран Западной Европы, где в лучших охотничьих хозяйствах достигается плотность до 36 птиц на 1 км2 (Шотландия; Moss et al., 1978) и где лицензионная стоимость отстрела одного глухаря (осень) составляет 100 фунтов (в ценах 1978 г.).
Основными биотехническими мероприятиями по сохранению и увеличению численности глухаря являются обеспечение охраны размножающихся птиц от хищников и факторов беспокойства и зимняя подкормка. Охрана от хищников особенно необходима в сравнительно изолированных лесных массивах, окруженных густонаселенной местностью, поскольку именно в таких ситуациях численность многих хищников (лисиц, ворон, одичавших собак и кошек) заметно выше, чем в естественных условиях. Например, владельцы высокорентабельных охотничьих ферм в Шотландии ведут систематическое истребление лисиц и ворон. Фактором беспокойства является прежде всего человек, и поэтому необходимо всячески избегать любых посещений им мест размножения в репродуктивный сезон. Именно по этой причине в большинстве охотничьих хозяйств в Западной Европы разрешается только осенняя охота на глухарей (в Пиренеях, например, с 24 сентября по 15 октября, по субботам, воскресеньям и средам, всего 10 дней в году; Gindre, 1978).
В условиях таежной зоны СССР весенняя охота пока еще вполне возможна, но для избежания отрицательных моментов (отстрел производителей, фактор беспокойства) необходимо соблюдать три обязательных условия.
1. Охотиться на токах только после завершения кульминационного момента токования — спаривания, занимающего около 10 дней. После прекращения появления самок на токовище самцы токуют здесь еще н.е менее двух недель, и в это время возможно проведение охоты.
2. Отстреливать на таких, как правило, небольших токовищах не более 1/3 взрослых самцов.
3. В хозяйствах с весенней охотой на глухаря иметь одно или несколько резервных токовищ, доступ на которые вплоть до конца июля должен быть категорически запрещен. Последнее условие непременно, поскольку весенняя стрельба на токах в любых случаях приводит к их деградации (Островский, 1974; Романов, 1979; мои данные).
Подкормка глухарей была введена в практику сравнительно недавно благодаря успешным опытам охотоведа И.И. Скутина, преследовавшего поначалу совсем иные цели (привлечение глухарей и тетеревов в глубь лесных массивов во избежание отравления гербицидами на полях). Подкормка проводится в местах обитания глухарей на специальных крытых платформах преимущественно зерновыми культурами (снопы необмолоченной пшеницы и овса) в зимнее время (Борискин, 1974). Такая подкормка оказалась исключительно полезной для-самок в ,период, предшествующий откладке яиц. Применение этого метода в охотхозяйствах Свердловской обл. уже привело к увеличению численности глухарей в 2.5—3 раза (Губин, 1981).
Охрана вида по всей территории ареала требует конкретных мероприятий прежде всего в местах интенсивных лесозаготовок и сплошной мелиорации.
Как хорошо известно, сплошная вырубка леса на токовище прекращает функционирование последнего и вызывает быстрое сокращение численности местных популяций. Как было показано выше, на большей части ареала вида токовища располагаются обычно в сосняках невысокого, IV—V классов, бонитета, обычно по окраинам верховых болот, и изъятие таких участков из лесозаготовительных планов не наносит последним ущерба. Охрана токовищ в данной ситуации тесно соприкасается с проблемами охраны верховых болот, встающими все более остро. В настоящее время Госкомитет по Лесному хозяйству СССР утвердил (в 1980 г.) инструкцию о защитных лесах, где запрещается рубка леса радиусом в 300 м вокруг токовищ.
Существенное значение имеет работа по восстановлению ареала вида, реинтродукции в места, где глухарь исчез уже в историческое время. В пределах СССР успех был достигнут при реинтродукции глухаря в сосняках в районе Борового, Кокчетавской обл. Здесь в 1965—1968 гг. было выпущено 4 партии взрослых птиц общим количеством в 135 особей. В 1980 г. здесь уже насчитывалось 699 глухарей (Черепанов, 1981). Для проведения такого рода работ большое значение имеет разведение глухарей в неволе. В последнее время здесь, был достигнут заметный прогресс (Немцев и др., 1973; Кутовая, 1976).
Географическая изменчивость. Занимая обширные территории, подчас весьма различные по условиям, этот практически оседлый вид демонстрирует значительную географическую изменчивость, хотя сплошное распространение и подвижность части популяций (особенно молодых особей) препятствуют выработке четких географических форм, выявлению которых мешает также индивидуальная и возрастная изменчивость. Последняя особенно, существенна, поскольку на коллекционных экземплярах нет практической возможности точно определять возраст самцов старше одного года, а многих молодых самок невозможно отличить от старых уже в первую зиму. Трудности же учета возрастной изменчивости могут приводить к ошибкам в оценке степени географической изменчивости. Все это, однако, не мешает выделить в настоящее время 12 явно выраженных географических форм.
Большинство подвидов глухаря, описанных к настоящему времени, далеко не равнозначны по своему таксономическому весу, и большой разнобой в трактовке отдельных форм глухарей разными авторами хорошо это иллюстрирует.
Наиболее интересна трактовка внутривидовой изменчивости глухаря, даваемая Иогансеном (Johansen, 1957). Он предполагал однородность популяций, населяющих одну и ту же природную зону и дающих в ее пределах клинальную изменчивость. Согласно этому автору, самая северная полоса таежной зоны оказывается населенной одним подвидом. Более южная полоса смешанных лесов населена все же разными подвидами, но сходными между собой и объединяемыми в одну группу. И, наконец, особо выделяется группа юго-западных подвидов. Эта схема вполне логична, но в действительности такой картины не существует. Неодинаковость природных условий в пределах одной и той же зоны в сочетании со сложной историей расселения и эволюции вида дает в результате весьма непростую картину, которую невозможно уложить в прокрустово ложе зональных границ.
Предпринятая мною ревизия показывает, что северная полоса таежной зоны населена по крайней мере тремя хорошо отличимыми друг от друга формами — западной номинативной, населяющей север Скандинавии и живущей в условиях довольно влажного морского климата; Кольской (Т.u. lonnbergi), обитающей на Кольском п-ове, природные условия которого представляют сложное сочетание суровых условий севера и теплого влияния Гольфстрима и, наконец, крупной и тускло окрашенной Т.u. obsoletus, занимающей север Восточной Европы и Сибири и живущей в условиях холодного и континентального климата. Большую часть континентальной Западной Европы и юг Швеции населяет наиболее темный и длиннохвостый Т.u. major, для всей области обитания которого характерна сравнительно мягкая и недолгая зима, даже в горах. Самые южные островки бореальных лесов, изолированные в горных поднятиях Кантабрии, Пиренеев, Восточных Карпат, занимают темные и сравнительно длиннохвостые подвиды Т.u. cantabricus, Т.u. aquitanicus, Т.u. rudolfi. В центральной части Восточной Европы обитает Т.u. pleskei — раса, по всем признакам переходная от Т.u. major к Т.u. obsoletus я к Т.u. volgensis. Последняя, населяющая бассейн средней Волги, весьма близка Т.u. uralensis, занимающей наиболее континентальные части южной части ареала, от Южного Урала до предгорьев Алтая, но формы эти тем не менее вполне отличимы одна от другой и сводить их в синонимы, на мой взгляд, нельзя. Юго-восточную часть ареала населяет Т.u. taczanowskii, отличающийся, как и западноевропейские подвиды, очень крупными размерами и сравнительно длинным хвостом при наличии значительно более светлой и струйчатой окраски.
Подвидовая диагностика глухарей базируется на признаках, наименее подверженных индивидуальной изменчивости. Это как особенности окраски (развитие коричневого цвета на крыле и на спине, струйчатого рисунка на различных частях оперения, в особенности на перьях груди; окраска шеи и поясницы; наличие и величина белых вершинных полос на НКХ и ВКХ и на РП), так и особенности морфологии (относительная длина хвоста) и этологйи (особенности брачной песни). В определенной степени учитываются также размеры и масса птиц, хотя они подвержены значительным индивидуальным колебаниям.
Проведенная мною ревизия позволяет следующим образом сгруппировать подвиды глухаря:
а) группа светлых континентальных подвидов — Т.u. urаlensis, Т.u. taczanowskii, Т.u. volgensis, Т.u. karelicus;
б) группы крупных, сравнительно длиннохвостых подвидов, более темноокрашенных, с меньшим развитием струйчатого рисунка и примитивным типом песни — североевропейские и северосибирские Т.u. obsoletus и Т.u. pleskei, связанные с предыдущей группой рядом популяций переходного типа;
в) темные, длиннохвостые, крупные с усложненным типом песни и минимальным развитием струйчатости подвиды Западной Европы — Т.u. urogallus, Т.u. major, Т.u. aquitanicus, Т.u. cantabricus;
г) горные дериваты западноевропейских форм, но с примитивным типом песни; наиболее северная — Т.u. lonnbergi и наиболее южная — Т.u. rudolfi.
Литература
• Фауна СССР. Академия наук СССР. Зоологический институт. Новая серия № 133. Птицы. Том III. Вып. 1. Р.Л. Потапов. Отряд Курообразные (Galliformes). Часть 2. Семейство Тетеревинные (Tetraonidae). Ленинград. Издательство «Наука». Ленинградское отделение. 1985 г. стр. 293-340.
Литература
• Коблик Е.А. 2025. Фауна птиц стран Северной Евразии в границах бывшего СССР: Списки видов.
• IOC World Bird List v15.1 (Updated February 20, 2025) by Frank Gill, David Donsker & Pamela Rasmussen.
• Н.Н. Балацкий. Птицы Северной Евразии. Таксономический список: август 2023 г.
• «Птицы СССР. Курообразные. Журавлеобразные». т.2. Р.Л. Беме, Н.П. Грачев, Ю.А. Исаков, А.И. Кошелев, E.Н. Курочкин, Р.Л. Потапов, А.К. Рустамов, В.Е. Флинт. Ленинград. Издательство «Наука». 1987 г. стр. 165-186.
Сайты
• zmmu.msu.ru
• www.balatsky.ru
• www.worldbirdnames.org
• Avibase by Denis Lepage
• Zoonomen by Alan Peterson
• Taxonomy by John Boyd
• сайт Василия Вишневского
• www.xeno-canto.org
• www.featherbase.info, www.pbase.com, www.macaulaylibrary.org