

Статус. Тростниковая сутора Paradoxornis heudei (David, 1872) является редким, стенотопным и локально распространённым гнездящимся, кочующим и зимующим видом. В Приморском крае он является сравнительно недавним вселенцем и представлен подвидом P. h. polivanovi Stepanyan, 1974) (рис. 1), внесённым в Красные книги Российской Федерации (2021) и Приморского края (2005).
Замечания по систематике. Во многих русских литературных источниках форма polivanovi, описанная Л.С. Степаняном (1974) по материалам, собранным на озере Ханка в 1971-1972 годах, считается самостоятельным видом (Степанян 1998, 2003; Коблик и др. 2006; Нечаев, Гамова 2009; Коблик, Архипов 2014), к которому причислили и подвид mongolicus из Восточной Монголии (Степанян 1998). Согласно нашим исследованиям, проведённым в Приморском крае (Глущенко, Шибнев 1981; Глущенко и др. 1995, 2016а,б) и Восточном Китае (Глущенко, Парилов 2000; Глущенко и др. 2010, 2012), придание видового статуса этим двум подвидам, объединённым в группу (allospecies) polivanovi-mongolicus, не оправдано, что уже дважды было достаточно подробно изложено ранее (Глущенко и др. 2016а,б).
В зарубежной орнитологической литературе идея разделения тростниковой суторы на два вида также не была поддержана многими исследователями (MacKinnon, Phillipps 2000; Robson 2007; Gombobaatar 2009; Del Hoyo, Collar 2016; The CNG Field Guide… 2024; и др.) и лишь некоторые авторы, например, Марк Брэзил (Brazil 2009), с ней согласились, хотя в последнем случае и ареал этих «видов», и особенности их внешней морфологии, были значительно искажены (Глущенко и др. 2016а).
Распространение и численность. Впервые на территории России тростниковую сутору обнаружили в южной части озера Ханка в 1968 году (Поливанов и др. 1973), а позднее этих птиц наблюдали и во многих

других районах Приханкайской низменности (Поливанова и др. 1974; Глущенко, Шибнев 1981; Назаров, Куринный 1981; Глущенко и др. 1995; и др.). Общая численность ханкайской гнездовой группировки, обитающей в российском секторе Приханкайской низменности (безусловно, самая крупная группировка в Приморском крае) для разных лет оценивалась в 400 (Глущенко, Шибнев 1981) и 230 (Глущенко и др. 1995) гнездящихся пар. По приблизительным расчётам, в пределах государственного природного биосферного заповедника «Ханкайский» обитает около 30% ханкайской популяции вида, и примерно такое же число птиц населяет его охранную зону (Лебяжинская, Глущенко 1995).
В окрестностях Уссурийска тростниковых сутор впервые встретили 26 октября 1982, когда группу из 3 особей наблюдали в долине реки Комаровка (Глущенко и др. 1986), а в 2002-2023 годах этих птиц периодически регистрировали в разных местах пригорода и находили их гнёзда (Глущенко и др. 2006а, 2019). К северу от Уссурийска кочующую тростниковую сутору (вероятно, отбившуюся от стаи) Е.А.Волковская-Курдюкова наблюдала в окрестностях села Михайловка (Михайловский район) 26 сентября 1998 (Глущенко и др. 2001).
В дельте реки Раздольная тростниковых сутор впервые отметили 22 мая 1983 (Глущенко и др. 1986), затем их встретили здесь 25-28 октября 1986 (Назаров 2004), а в 1998 году их нашли достоверно гнездящимися (Нечаев, Горчаков 2001). С тех пор они там регулярно размножаются, а их численность, по нашим экспертным оценкам, составляет 30-50 пар и зависит от степени сохранности густых зарослей тростника, подверженных регулярному выжиганию (Глущенко и др. 2016а).
Западнее устья реки Раздольная тростниковые суторы локально заселили северное побережье Амурского залива, в том числе, они были найдены гнездящимися в районе устья реки Шмидтовка (Омелько, Омелько 2010; Сотников и др. 2016; Глущенко и др. 2016б), а также в окрестностях села Прохладное, станции Амурский Залив и посёлка Угловое (Глущенко и др. 2016а). Помимо этого, птиц отмечали у побережья Уссурийского залива в приустьевых частях рек Артёмовка и Шкотовка, а также на побережье озера Кролевецкое (Глущенко и др. 2016а).
К югу от устья Раздольной, в низовье реки Пойма, мы впервые наблюдали тростниковых сутор в 2002 году. В 2014 году этих птиц встречали в районе лагуны Цапличья, в бухте Бойсмана, а также на ручье, впадающем в бухту Северная, где 31 мая 2016 года обнаружили гнездо.
На крайнем юге Хасанского района (урочище Туманган) в 2014 и 2015 годах тростниковых сутор регистрировали в районе залива Голубиный и бухты Сивучья (Глущенко, Коробов 2014; Сотников и др. 2016), а в мае-июне 2016 года в окрестностях озера Лотос (Дорицени) здесь впервые нашли их гнёзда. В настоящее время в подходящих биотопах они локально гнездятся на всех заболоченных участках от залива Посьет на севере до низовий реки Туманная на юге.
Во внутренних районах Приморского края в июле 1984 года тростниковых сутор обнаружили в среднем течении реки Уссури в окрестностях города Арсеньев, где в небольшом количестве и в разные фенологические периоды их регулярно наблюдали (в том числе, находили гнездо и видели слётков) вплоть до 1989 года, но позднее, несмотря на специальные поиски, проводимые в 1990-1998 годах, этих птиц регистрировать не удавалось (Назаренко 2016).
Вероятно, локальные поселения тростниковых сутор могут временно возникать в поймах реки Сунгача и в низовьях реки Уссури, а в бассейне нижнего течения реки Бикин в зимний и летний периоды этих птиц наблюдали на Лучегорском водохранилище (Бурковский 1996; 1998). Таким образом, к настоящему времени тростниковая сутора широко, хотя и крайне локально, заселила значительную часть равнинных и прибрежных безлесных участков западного сегмента Приморского края (рис. 2).
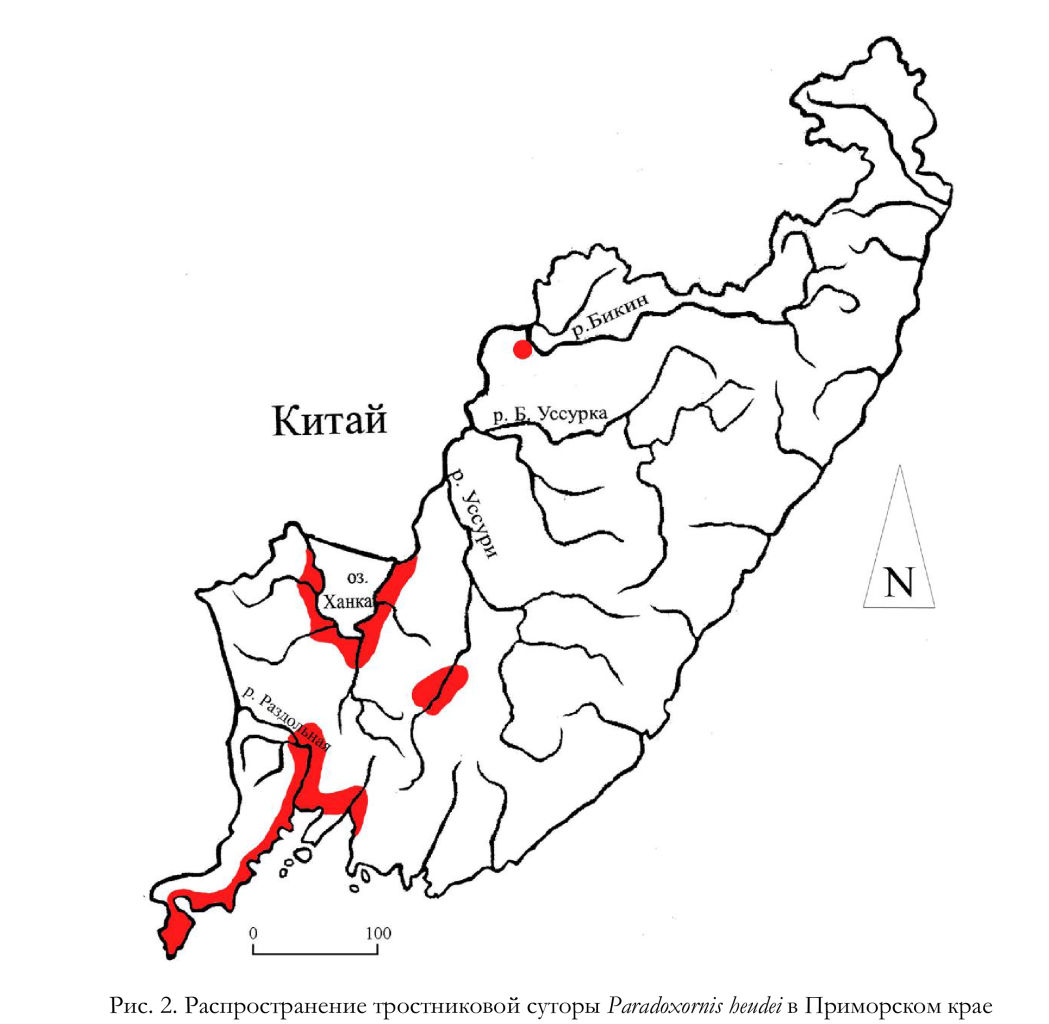
Согласно нашей экспертной оценке, на начало 2020-х годов в Приморском крае гнездилось 250-500 пар тростниковых сутор. При этом существуют значительные колебания численности на отдельных участках, а также происходит активная территориальная экспансия. Этот процесс выглядит не как расселение вида на фоне роста численности в основном (ханкайском) гнездовом анклаве, а как вынужденное выселение части птиц за его пределы. Одной из причин такой динамики в последние годы можно считать значительное ухудшение условий обитания на Приханкайской низменности, которое обусловлено очень высоким уровнем воды в озере Ханка. В связи с этим ёмкость гнездовых угодий тростниковых сутор уменьшилась, и гнездившаяся группировка птиц была вынуждена распределяться по смежным территориям (Глущенко и др. 2016а).

Изучение истории находок тростниковой суторы в Приморском крае позволяет предположить, что этот вид для Приморья является сравнительно недавним вселенцем (Глущенко и др. 2016а; Назаренко 2016).
Предположительно, эти птицы впервые проникли на Приханкайскую низменность в начале второй половины ХХ столетия, впоследствии продолжив расселяться в разных направлениях, в частности на юг, дойдя к настоящему времени до крайней южной оконечности Приморья (Глущенко и др. 2016а). Следует отметить, что имевший место в последней четверти ХХ века упадок сельского хозяйства в Приморском крае мог положительно сказаться на успехе экспансии тростниковой суторы, поскольку многие сельскохозяйственные земли (в частности, расположенные на Ханкайско-Раздольненской равнине) превратились в многолетние залежи, отчасти зарастающие тростником. Даже если в большинстве локусов эти заросли не могли служить гнездовым биотопом данного вида, они способствовали выживанию птиц во время осенне-зимних кочёвок, создавая экологические коридоры, в которых они могут некоторое время кормиться и скрываться от хищников. Это обстоятельство очень важно, учитывая сравнительно слабые лётные качества тростниковых сутор (Глущенко и др. 2016а).

Местообитания. Во все сезоны годового цикла тростниковые суторы строго приурочены к тростниковым зарослям (рис. 3). В типичном случае в гнездовой период они явно избегают селиться в изреженных, изолированных и низкорослых зарослях этого растения, а также на участках с хорошо выраженным рельефом либо расположенных в относительно сухих местах. В холодную часть года тростниковые суторы кочуют, надолго задерживаясь только среди тростниковых массивов (рис. 4).
Гнездование. По мнению Н.Н. Поливановой с соавторами (1980, 2009), у тростниковых сутор пары сохраняются надолго и формируются в различных комбинациях из старых и молодых особей, что, вероятно, происходит осенью. Согласно другим наблюдениям, формирование пар может происходить во второй половине марта (Глущенко, Шибнев 1981).
В 1972 году на озере Ханка к постройке гнёзд суторы приступили с начала третьей декады мая. Первую самку со строительным материалом здесь встретили 16 мая, а массовое гнездостроение наблюдали с 19 по 31 мая (Поливанова и др. 1980, 2009).
По данным В.А. Нечаева и Г.А. Горчакова (2001), в устье Раздольной тростниковые суторы приступали к гнездованию в мае, причём сроки начала постройки гнёзд зависели от защитных условий тростниковых зарослей, при этом птицы не начинали строить гнёзда до тех пор, пока побеги молодого тростника не вырастут на 50-70 см.
По нашим данным, в настоящее время к строительству первых гнёзд суторы иногда приступают в начале мая, а в отдельных случаях даже в последней декаде апреля. Подтверждением этого является находка на северном побережье Амурского залива, в приустьевой части реки Шмидтовка, гнезда с полной кладкой из 4 яиц уже 3 мая 2021, таким образом, согласно расчётам, начало строительства гнезда в этом случае приходилось на период с 22 по 26 апреля.
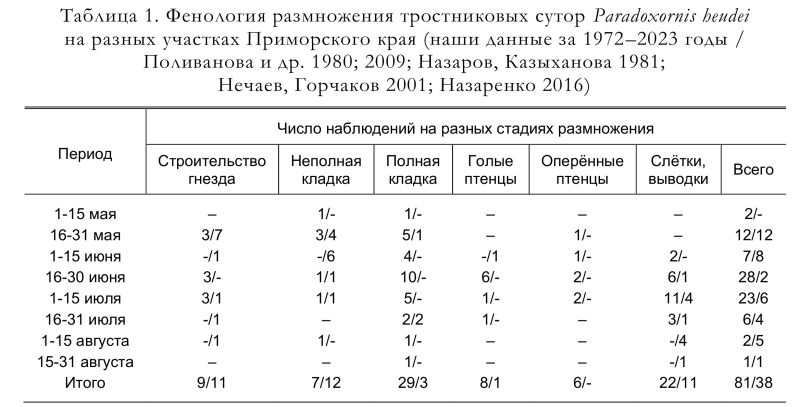
В связи с сильными ветрами, часто разрушающими гнёзда, построенные в недостаточно густых зарослях тростника, гнездостроение многих пар может сильно затягиваться. В целом, учитывая этот факт, а также наличие вторых, и, вероятно, третьих циклов размножения успешно размножавшихся пар (Назаров, Казыханова 1981; Глущенко и др. 2016б), в Приморье гнездовой период тростниковых сутор растянут по меньшей мере на четыре месяца – с начала мая до конца августа (табл. 1). Более того, встреча молодых птиц с недоросшими рулевыми перьями в начале октября (Глущенко, Шибнев 1981) позволяет предполагать, что в отдельных случаях размножение может быть ещё более поздним.
По данным Н.Н. Поливановой с соавторами (1980, 2009), на озере Ханка тростниковые суторы как правило устраивали гнёзда в куртинах тростника, заметно отличающихся от окружающих зарослей большей высотой, густотой и желтизной, а также прямыми несломленными ветром стеблями. Такие куртины обычно окружены плёсами или редкими тростниками. Площадь их варьирует от 50 до 100 м2. Внутри куртин нередки участки воды, лишённые растений или с небольшими группами тростника разной густоты. В сплошных зарослях тростника птицы гнездятся реже (2 гнезда из 12 найденных) (Поливанова и др. 1980, 2009).
Мы также отметили тот факт, что многие гнёзда находились вблизи прогалов в тростниковых крепях в виде проток, узких троп или значительно разреженных участков.
За очень редким исключением, тростниковые суторы размещают свои гнёзда на стеблях тростника (рис. 5). Лишь одно из 52 найденных нами гнёзд было устроено на стеблях полыни (Глущенко и др. 2016а; наши данные), а ещё одно крепилось к стеблям тростника и спиреи иволистной Spiraеa salicifolia.
Обычно гнёзда располагаются в густых зарослях тростника и прикрепляются к 2-8 (чаще всего к 4-6) вертикальным стеблям. Реже птицы строят их в относительно разреженных и низких тростниках (при наличии толстых стеблей, вершины которых сломаны), а ранние постройки могут быть скрыты под заломами тростниковых стеблей (Глущенко и др. 2016а; рис. 5.7).
Высказывалось мнение, что если один из партнёров гнездовой пары является старой особью, то в случае гибели гнезда птицы начинают строить новое на освоенном участке, не далее 10-15 м от места расположения прежнего гнезда, а если пара состоит из двух молодых птиц, то после потери гнезда они перемещаются дальше (Поливанова и др. 1980, 2009). По нашим сведениям, безотносительно возраста половых партнёров новые гнёзда нередко располагаются неподалёку (от 7 до 20 м) от старых, а 22 мая 2017 у восточного побережья озера Ханка свежевыстроенное гнездо птицы разместили под прошлогодним гнездом и на тех же тростинках, при этом расстояние от низа старой до верха новой постройки составило всего 54 мм.

По данным Ю.Н. Назарова и М.Г. Казыхановой (1981), у восточного побережья озера Ханка высота расположения гнёзд тростниковых сутор варьировала от 28 до 103 см, в среднем составляя 61 см. По другим сведениям, гнёзда устраиваются у основания стеблей тростника на высоте от 18 до 76 см над водой, чаще в 30-50 см. К моменту его постройки зелёные стебли тростника поднимаются над водой на 60-100 см и достигают приблизительно одной трети высоты прошлогодних (Поливанова и др. 1980, 2009). Согласно нашим наблюдениям, птицы строили гнёзда на высоте от 30 до 160, в среднем около 65 см (n = 22) над водой, сплавиной или почвой. Известно, что в первой половине лета гнёзда устраиваются заметно ниже, чем во второй (Назаров, Казыханова 1981; наши данные).
Начиная строить гнездо, птицы соединяют несколько стеблей тростника клочками свалявшейся паутины, а затем вплетают полоски основного материала; строительство гнезда идёт сверху вниз, при этом гнёзда могут крепиться к опорным стеблям разным образом: только верхним краем, верхним и нижним краями, серединой, или по всей высоте (Назаров, Казыханова 1981).
В первой половине лета птицы строили гнездо в течение 3-5 дней, при этом постройки располагались в густом переплетении сухого тростника, образующего кочку среди воды, а во второй половине лета они устраивали гнёзда на окраине более или менее густых зарослей тростника, маскируя их сверху, надламывая стебли, к которым они крепятся, формируя своеобразный навес в 40-50 см от верхнего края постройки (Назаров, Казыханова 1981). По другим данным (Поливанова и др. 1980, 2009), в тихую погоду на строительство гнезда затрачивается 3-4 дня, а в ветреную – до 8 дней. Пары, бывшие под наблюдением, занимались строительством гнезда в течение всего дня, однако более интенсивно работали в его первой половине. 24 мая с 7 до 11 ч самка прилетала со строительным материалом 45 раз, а с 17 до 20 ч – 20-25 раз; собирала она его в 2-10 м от гнезда. Птицы летали за материалом по трём направлениям: 75 % случаев по одному, в 15 % – по второму, в 10 % – по третьему (Поливанова и др. 1980, 2009).
Н.Н. Поливанова с соавторами (1980, 2009) указывает, что единственным материалом, который тростниковые суторы используют для строительства гнезда, служат стебли тростника, при этом самки предварительно расщепляют их клювом на узкие продольные полоски разной длины, что требует много времени и спокойной обстановки. Дно и стенки гнезда строят из мягких и тонких оболочек тростникового стебля, расщеплённого на узкие (до 5 мм) и короткие (до 5 см) полоски, укладывая их в вертикальном направлении или по диагонали, реже горизонтально.
При строительстве основного каркаса гнезда птицы не вплетают стебли тростника в его стенку по всей высоте, а прикрепляют гнездо к стеблям лишь в некоторых местах, обматывая их волокнами, концы которых вплетают в стенки гнезда. Дно постройки составляет около 1/3 высоты всего гнезда. Верхний край создаётся из вершинных частей стеблей тростника, которые укладываются кольцеобразно. Иногда птицы вплетают в него расщеплённый вдоль мягкий эластичный пористый стержень тростника. Полоски строительного материала укладываются аккуратно одна к другой, перемежаясь с узкими мягкими плёнками внешней оболочки стеблей. Этим же материалом птица тщательно прикрепляет гнездо к стеблям. Весь строительный материал обильно скрепляется паутиной. Лоток создаётся быстро, из верхушечных частей стебля. При строительстве лотка самке активно помогает самец, который садится в гнездо, формует и подправляет лоток (Поливанова и др. 1980, 2009).
По данным Ю.Н. Назарова и М.Г. Казыхановой (1981), у восточного побережья озера Ханка материалом для строительства гнёзд служили полоски (шириной 3-5 мм) влагалищ листьев тростника, плотно скрепляемые паутиной; снаружи птицы облицовывали их некоторым количеством узких (около 1 мм) тонких полосок стеблей тростника, что иногда придавало гнезду неряшливый вид, так как его стенки казались рыхлыми; лоток выстилался узкими полосками стеблей, а в одном случае в нём было небольшое количество узких полосок зелёных листьев тростника. Строительный материал одного из гнёзд, найденного в устье реки Раздольная 13 июля 1999, на 95% состоял из узких полосок от сухих листовых влагалищ, прикрывающих стебель тростника; эти полоски переплетались между собой как в горизонтальном, так и в вертикальном положениях; реже птицы использовали узкие полоски от стеблей (соломинки) и листьев тростника; на дне гнезда находились стеблевые волокна, которые были согнуты или торчали в беспорядке. Снаружи гнездовой материал был облицован тонким слоем паутины, которая покрывала и края лотка, оплетая сверху стеблевые полоски, а в лотке были более узкие и тонкие волокна листовых влагалищ тростника (Нечаев, Горчаков 1981).
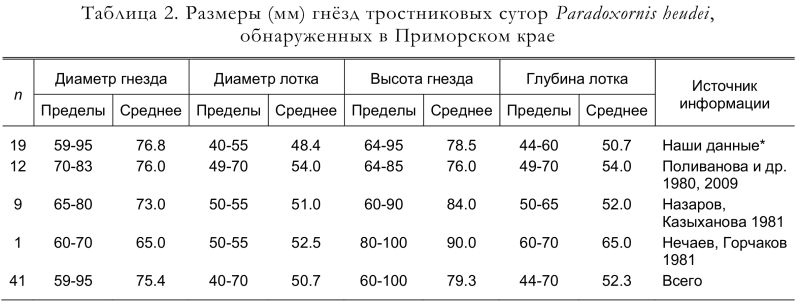
Размеры гнёзд приведены в таблице 2. Отметим, что гнёзда имеют узкий и глубокий лоток, что предотвращает выпадение из них птенцов во время сильных ветров, но, по-видимому, может служить причиной гибели младших птенцов, затаптываемых более крупными и сильными старшими (Глущенко, Шибнев 1981).
От момента начала постройки гнезда до откладки первого яйца проходит 4-8 дней; в этот промежуток времени обе птицы держатся на своём участке, часто посещая гнездо (Назаров, Казыханова 1981).
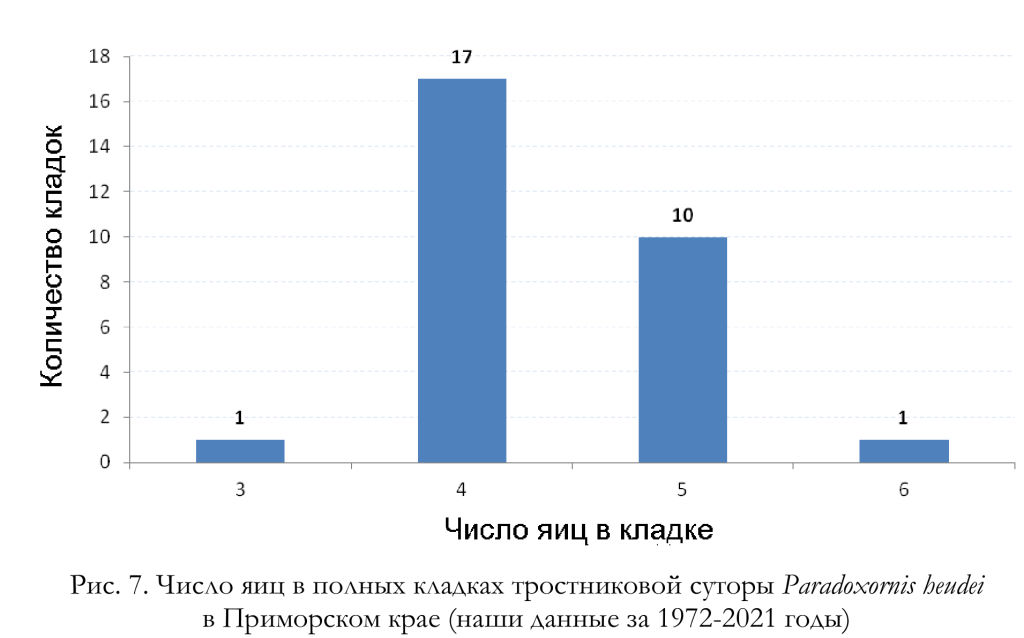
По данным Н.Н. Поливановой с соавторами (1980, 2009), из 10 кладок, находившихся под наблюдением, 8 содержали 5 яиц, 1 – 4 и 1 – 6 яиц (то есть в среднем на гнездо приходилось 5.0 яйца). Согласно Ю.Н. Назарову и М.Г. Казыхановой (1981), полные кладки тростниковых сутор у восточного побережья озера Ханка в 1977-1978 годах содержали от 2 до 5, в среднем 3.9 яйца. По нашим материалам, полные кладки включают от 3 до 6, в среднем 4.38 яйца (n = 29) (рис. 6, 7).


Линейные размеры, индекс удлинённости и объём яиц тростниковой суторы приведены в таблице 3. Согласно нашим материалам, вес свежих и слабо насиженных яиц варьировал от 1.6 до 2.5 г, составляя в среднем 1.92 г (n = 31). Объём яиц* находился в пределах 1.3-2.3 см3, составляя в среднем 1.78 см3 (n = 76).
По данным Ю.Н. Назарова и М.Г. Казыхановой (1981), окраска яиц несколько варьирует, при этом основной фон белый (обычно со слабым кремовым оттенком); по всей поверхности разбросано сравнительно небольшое количество глубинных и поверхностных бледно-оливковых, коричневатых, светло- или темно-бурых мазков, которые сгущаются вокруг тупого конца, а иногда мазки перемежаются мелкими крапинками такого же цвета. Согласно описанию Н.Н.Поливановой с соавторами (1980, 2009), фон яиц белый с крапом из светло-зелёных и светло-коричневых пятен, при этом крапинки сгущаются в средней части яйца, где обычно образуется венчик у тупого полюса. Окраску яиц некоторых найденных нами кладок иллюстрируют рисунки 6 и 8


Н.Н. Поливанова с соавторами (1980, 2009) указывают, что насиживание продолжается 13 дней. Кладку насиживают оба родителя, регулярно меняясь. В одном из гнёзд на 10-й день насиживания за светлое время суток птицы обогревали яйца 13 ч 40 мин, при этом самка – 9 ч 33 мин, а самец – 4 ч 07 мин. Ночью на гнезде оставалась самка. Во время насиживания она сидела в гнезде глубже и при взгляде сбоку были видны только клюв и хвост, напоминающий листок тростника. У самца, сидящего на гнезде, видна верхняя часть головы. Партнёры менялись через 30-60 мин, чаще в утренние часы и реже днём. Насиживающая птица покидает гнездо за 20-30 с до прилёта сменяющей её особи, при этом смена партнёров происходит молча (Поливанова и др. 1980, 2009). По другим данным, насиживание начинается ещё до завершения кладки, поэтому вылупление птенцов растягивается на 2 сут, а птенцы появляются через 12-14 сут после окончания кладки (Назаров, Казыханова 1981). Насиживающая птица сидит очень плотно (рис. 9), обычно подпуская наблюдателя на расстояние менее 1 м (Назаров, Казыханова 1981).


Согласно материалам, собранным Н.Н.Поливановой с соавторами (1980, 2009), птенцы появляются на 14-й день насиживания. Режим их обогревания в день вылупления почти не отличается от такового при насиживании кладки в последние дни. Так, в одном из гнёзд за светлое время суток самка провела на только что вылупившихся птенцах 9 ч 21 мин, а самец – 4 ч 56 мин. В целом гнездовые птенцы (рис. 10) обогревались родителями 14 ч 17 мин. Ночью так же, как и во время насиживания, на гнезде оставалась самка. Вылупившихся птенцов родители (главным образом самец) сразу же начинают кормить. Шестидневных птенцов взрослые кормили с 5 ч 05 мин до 20 ч 25 мин, принеся им пищу 112 раз (самец 63 раза, самка – 49). Десятидневных птенцов птицы кормили 136 раз – с 5 ч 11 мин до 20 ч 37 мин (самец 96 раз, самка – 40 раз).
Таким образом, главную роль в выкармливании птенцов играет самец. Он не только чаще прилетает к гнезду с пищей, но, как правило, приносит её больше, чем самка. Отправляясь на охоту, он дольше отсутствует и дальше улетает от гнезда. Самка приносит меньше корма, собирает его поблизости и быстро возвращается в гнездо, так как функция обогревания птенцов (рис. 11.1,2) лежит в основном на ней (Поливанова и др. 1980, 2009).
Только что вылупившиеся птенцы весят от 1.3 до 1.4 г, а к концу первого дня их вес увеличивается до 1.65 г. Ввиду неодновременности вылупления птенцов отставание в развитии младшего из них сохраняется в течение всего времени пребывания в гнезде. Разница в весе младшего и старшего птенцов доходит до 1.5 г. К концу периода пребывания в гнезде вес птенцов увеличивается почти в 10 раз (Поливанова и др. 1980, 2009).
В гнёздах мы находили от 1 до 5 птенцов, а в среднем на одно гнездо приходилось 3.47 птенца (n = 15). В их выкармливании (рис. 11.3) принимали участие оба партнёра.

Птенцы покидают гнёзда на 11-12-й день, при этом, выпрыгивая из гнезда, они сразу же усаживаются на соседних стеблях тростника не далее 50-70 см от него и цепко держатся за стебли, пытаясь взбираться по ним вверх, но быстро устают и перепрыгивают на горизонтально лежащий стебель. Первые дни после оставления гнезда они держатся возле него в нижнем ярусе тростниковых зарослей. Обычно слётки рассаживаются близко один от другого и часто перекликаются. Птенцы ночуют рядом, прижавшись друг к другу (Поливанова и др. 1980, 2009).
По другим данным, птенцы покидают гнездо в возрасте 10-12 суток (Глущенко, Шибнев 1981). При удачном стечении обстоятельств вылет птенцов первого выводка случается в первой либо второй половине июня (рис. 12), и родители могут вновь приступать к гнездованию.

В 1972 году в бывшие под наблюдением гнёзда было отложено 45 яиц, из которых вылупилось 24 птенца (53.3 %), а выросло и вылетело из гнёзд 13 птенцов, что составило 28.9 % от числа отложенных яиц (Поливанова и др. 1980, 2009).
Территориальные перемещения, зимовка. Некоторые группы молодых птиц первого выводка, ставших самостоятельными, начинают кочевать уже с конца первой декады июня (рис. 13), а позднее кочёвки продолжаются всю вторую половину лета, осень и зиму.

Начиная с декабря тростниковые суторы заметно сужают район кочёвок по сравнению с летне-осенним периодом (Глущенко, Шибнев 1981).
Зимой они кочуют небольшими группами, численностью от нескольких птиц до двух-трёх десятков особей, в среднем (по 64 группам) 7.1 особь на группу. В разные зимы на Приханкайской низменности среднее число птиц в группах менялось от 5.3 до 8.0 особи, причём, по нашему убеждению, сформированному при работе в Приморском крае и Восточном Китае, оно в значительной мере отражает плотность населения и благополучие размножения всей локальной популяции вида. К концу зимы численность птиц в этих группах сокращается за счёт гибели части особей (Глущенко и др. 2006б).
Зимой 1977/78 года для учёта численности мы обследовали около 70 % территории Приханкайской низменности, пригодной для зимнего пребывания тростниковых сутор, при этом обнаружили 52 группы общей численностью около 390 особей. Плотность их населения в типичных местообитаниях достигала 30 особей на 1 км2, а общая расчётная численность составила 600-700 особей (Глущенко, Шибнев 1981).

По данным В.А. Нечаева и Г.А. Горчакова (1981), в дельте реки Раздольная тростниковые суторы – оседлые птицы, обитающие в благоприятных местах в течение нескольких лет, покидая район гнездования в многоснежные и холодные зимы; большую часть года они держатся стаями численностью от 3-4 до 10-15 особей.
Питание. Согласно данным Н.Н. Поливановой с соавторами (1980, 2009), на озере Ханка тростниковые суторы кормятся как открытоживущими, так и скрытоживущими беспозвоночными. Из первых в тростниковых зарослях обильны мелкие пауки и моллюски, а также бабочкисовки Noctuidae. Именно они служили основой питания взрослых птиц весной и летом; ими же родители выкармливали потомство. В нескольких пробах, взятых у гнездовых птенцов, оказались в основном мелкие пауки, моллюски, а также брюшко самки совки. Во время наблюдений у гнёзд в клюве прилетающих к птенцам птиц, кроме пауков, часто отмечались небольшие гусеницы и личинки других беспозвоночных (Поливанова и др. 1980, 2009) На восточном побережье озера Ханка в 10 пробах корма птенцов оказались гусеницы (18 экз.), личинки мух (2 экз.), куколка бабочки и пауки (3 экз.), при этом большинство этого корма суторы добывали на стеблях тростника (Назаров, Казыханова 1981). Согласно визуальным наблюдениям, основной пищей тростниковым суторам в летний период служат насекомые и их личинки (рис. 14), которых они собирают как с поверхности тростника, так и из стеблей. Главными компонентами пищи, приносимой птенцам, были гусеницы, личинки пилильщиков, прямокрылые и пауки (Глущенко, Шибнев 1981). В холодную часть года основную пищу птиц составляют личинки, которых тростниковые суторы извлекают из стеблей тростника (рис. 15).
Неблагоприятные факторы, враги, гибель. В дельте Раздольной одним из основных факторов, оказывающих отрицательное влияние на размещение, успешное гнездование и зимовку тростниковых сутор, являются травяные палы, во время которых тростниковые заросли сгорают на значительных площадях (Нечаев, Горчаков 1981). Ю.Н.Назаров и М.Г. Казыханова (1981) считают, что непременным условием охраны этого вида является предотвращение выжигания зарослей тростника, при котором сокращается пригодная для гнездования площадь и значительно снижается успешность размножения этих птиц.
По нашим данным, на многих участках обитания в Приморье эти птицы находятся в крайне сложной ситуации, вызванной регулярными травяными палами (рис. 16), уничтожающими основную часть тростниковых массивов. Например, на Приханкайской низменности в 1978-1980 годах травяные палы охватывали от 55 до 90% всего массива водно-болотных угодий (Глущенко, Бочарников 1989). По этой причине в окрестностях Уссурийска и в ряде других мест обитания тростниковых сутор в Приморье нет даже небольших по площади, но достаточных для размножения этого вида участков подходящих стаций, где он мог бы размножаться ежегодно (Глущенко и др. 2006а).
В недалёком прошлом происходило массированное осушение и распашка заболоченных территорий Приханкайской низменности, связанное главным образом с расширением зоны рисосеяния, а зимой 1972/73 года наблюдали выкашивание обширных массивов тростника на корм скоту, что значительно отразилось на численности птиц (Поливанова и др. 1980, 2009; Глущенко, Шибнев 1981).

К естественным негативным факторам можно отнести сильные ветры, разрушающие гнёзда сутор. Так, по данным Н.Н. Поливановой с соавторами (1980, 2009), построенные из относительно коротких и малоэластичных волокон тростника гнёзда плохо противостоят раскачиванию стеблей во время сильного ветра. Верхний край и остов гнезда, разнородные по укладке и структуре материала, постепенно теряют связь между собой и разделяются на две части. Петли тонких волокон, которыми верхний край гнезда прикрепляется к стеблям тростника, ослабевают и некоторые отрываются от постройки. В результате гнездо наклоняется, а потом и сваливается. Из 12 найденных гнёзд 5 погибло от сильных ветров. Особенно страдают гнёзда в начале их возведения. К качающимся стеблям тростника птица никак не может прикрепить строительный материал и бросает начатое гнездо. Если оно незакончено, рыхлое и плохо закреплено, ветер быстро разрывает и разносит сухие короткие волокна тростника, из которых оно свито. После гибели первой постройки суторы начинают возводить новую. Некоторые пары начинали строить гнёзда по 4-5 раз. Лишь у одной пары гнездостроение прошло благополучно от начала до конца. Гибель гнёзд от ветра затягивает сроки размножения. Как следствие этого, возникает ещё один неблагоприятный фактор – конкуренция со стороны восточных дроздовидных камышевок Acrocephalus orientalis. Обычно у них гнездостроение начинается позже, чем у тростниковых сутор, однако, когда сроки совпадают, восточные камышевки растаскивают строительный материал из незаконченных гнёзд тростниковых сутор (Поливанова и др. 1980, 2009).
Мы трижды находили гнёзда тростниковых сутор с расклёванными яйцами, при этом гнёзд других воробьиных птиц (помимо построек соседних пар тростниковых сутор) поблизости не было, что наводило на мысль о разорении этих кладок представителями своего же вида.
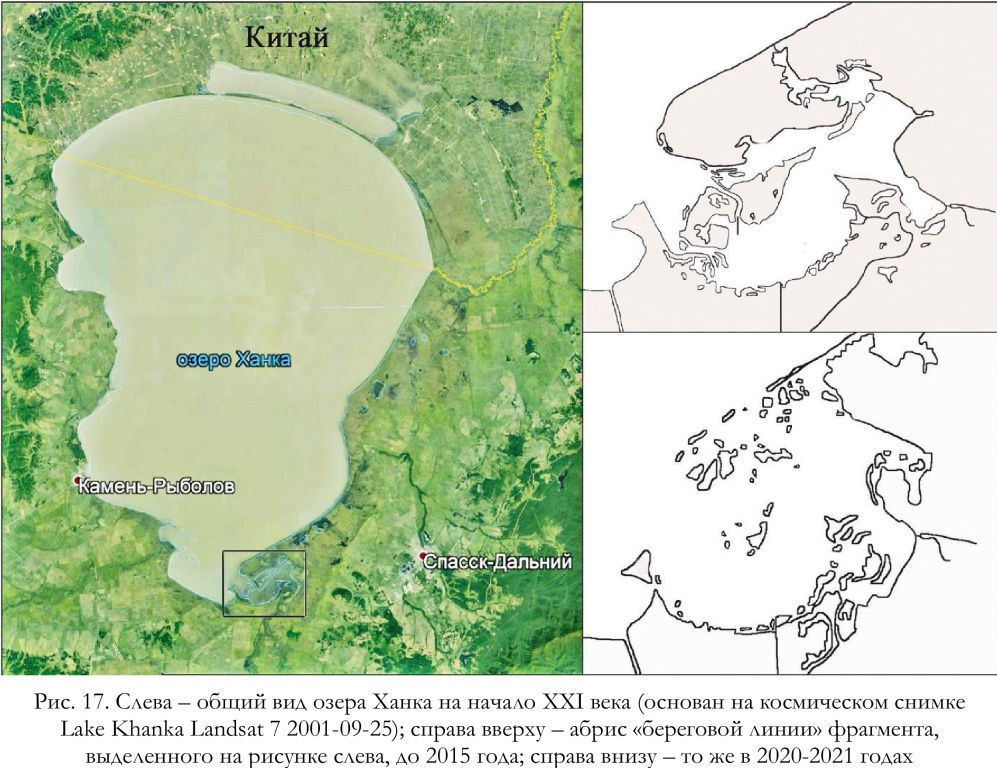

В 2015 году уровень воды в озере Ханка превысил исторический максимум (Бортин, Горчаков 2016), что привело к избыточному увлажнению и затоплению наиболее низких участков побережья и прилегающей к озеру котловины. В таких условиях значительные массивы тростника позднее оказались затопленными и разбитыми штормами (рис. 17, 18), став непригодными для обитания тростниковых сутор, что, безусловно, отрицательно сказалось на общей ёмкости гнездовых стаций (Глущенко и др. 2016а). О возможности негативного воздействия длительного подъёма уровня воды в озере Ханка, который вызывает разреживание и сокращение площади тростниковых крепей, и, соответственно, снижение численности местной популяции тростниковых сутор, упоминали ещё задолго до современного катастрофического положения, имеющего место в настоящее время (Поливанова и др. 1980).

Литература
• Бортин Н.Н., Горчаков А.М. 2016. Анализ факторов неустойчивости режима озера Ханка // Трансграничное озеро Ханка: причины повышения уровня воды и экологические угрозы. Владивосток: 31-40.
• Бурковский О.А. 1996. Встреча тростниковой суторы Paradoxornis heudei в северной части Приморского края // Рус. орнитол. журн. 5 (1): 6. EDN: RUZFLB.
• Бурковский О.А. 1998. Некоторые интересные встречи птиц в Приморье // Рус. орнитол. журн. 7 (43): 13-15. EDN: JRWZQN.
• Глущенко Ю.Н., Бочарников В.Н. 1989. Травяные пожары на Приханкайской низменности и рекомендации по борьбе с ними // Проблемы краеведения (Арсеньевские чтения). Ч. 2. Природное краеведение. Уссурийск: 64- 66.
• Глущенко Ю.Н., Волковская-Курдюкова Е.А., Коробов Д.В., Кальницкая И.Н. 2010. Материалы к изучению фауны и населения птиц китайского сектора бассейна озера Ханка // Амур. зоол. журн. 2, 4: 368-377.
• Глущенко Ю.H., Волковская-Курдюкова Е.А., Мрикот К.Н. (2001) 2015. Новые сведения о редких и малоизученных птицах Приморского края // Рус. орнитол. журн. 24 (1174): 2788-2792. EDN: UBSHAD.
• Глущенко Ю.Н., Коробов Д.В. 2014. Авифаунистические исследования на крайнем юго-западе Приморского края весной 2014 г. // Животный и растительный мир Дальнего Востока 2 (22): 6-14.
• Глущенко Ю.Н., Липатова Н.Н., Мартыненко А.Б. 2006а. Птицы города Уссурийска: фауна и динамика населения. Владивосток: 1-264.
• Глущенко Ю.Н., Ли Сяомин, Коробов Д.В., Волковская-Курдюкова Е.А., Кальницкая И.Н., Лю Хуа Цзинь, Ван Фэнкунь, Юй Веньтао. 2012. Учёты птиц в китайском национальном парке «Синкай-Ху» в 2011-2012 гг. и некоторые итоги авифаунистических исследований в бассейне озера Ханка // Амур. зоол. журн. 4, 3: 288-303.
• Глущенко Ю.Н., Коробов Д.В., Бурковский О.А., Тиунов И.М., Сурмач С.Г., Сотников В.Н., Акулинкин С.Ф. 2016. Тростниковая сутора Paradoxornis heudei в Приморском крае // Рус. орнитол. журн. 25 (1320): 2873-2884. EDN: WEFJNL.
• Глущенко Ю.Н., Коробов Д.В., Харченко В.А., Коробова И.Н., Глущенко В.П. 2019. Птицы – Aves // Природный комплекс Уссурийского городского округа; современное состояние. Владивосток: 151-301.
• Глущенко Ю.Н., Нечаев В.А., Редькин Я.А. 2016б. Птицы Приморского края: краткий аунистический обзор. М.: 1-523.
• Глущенко Ю.Н., Парилов М.П. 2000. Материалы по зимовке птиц в Восточном Китае // Проблемы экологии Верхнего Приамурья. Благовещенск, 5: 118-134.
• Глущенко Ю.Н., Шибаев Ю.В., Лебяжинская И.П. 1995. Современное состояние популяций некоторых редких видов птиц Приханкайской низменности // Проблемы сохранения водно-болотных угодий международного значения: Озеро Ханка (Тр. международ. науч.-практ. конф.). Спасск-Дальний: 45-50.
• Глущенко Ю.Н., Шибнев Ю.Б. 1981. Тростниковая сутора – Paradoxornis heudei David на озере Ханка // Редкие птицы Дальнего Востока. Владивосток: 56-63.
• Глущенко Ю.Н., Шибнев Ю.Б., Волковская-Курдюкова Е.А. 2006б. Птицы // Позвоночные животные заповедника «Ханкайский» и Приханкайской низменности. Владивосток: 77-233.
• Глущенко Ю.Н., Шибнев Ю.Б., Медведев В.Н. (1986) 2012. Заметки о новых для Приморья и редких птицах // Рус. орнитол. журн. 21 (810): 2693-2694. EDN: PENYDF.
• Коблик Е.А., Архипов В.Ю. 2014. Фауна птиц стран Северной Евразии в границах бывшего СССР: списки видов // Зоологические исследования 14: 1-171.
• Коблик Е.А., Редькин Я.А., Архипов В.Ю. 2006. Список птиц Российской Федерации. М.: 1-281.
• Лебяжинская И.П., Глущенко Ю.Н. 1995. Приоритетные направления развития природоохранной деятельности заповедника «Ханкайский» // Проблемы сохранения водно-болотных угодий международного значения: Озеро Ханка (Тр. международ. науч.-практ. конф.). Спасск-Дальний: 122-128.
• Назаренко А.А. 2016. Первое свидетельство расселения ханкайской популяции тростниковой суторы Paradoxornis heudei polivanovi Stepanyan, 1974 на прилежащие территории с комментариями о неоднозначной роли хозяйственной деятельности в биогеографической истории этого подвида // Дальневост. орнитол. журн. 5: 32-41.
• Назаров Ю.Н. 2004. Птицы города Владивостока и его окрестностей. Владивосток: 1-276.
• Назаров Ю.Н., Казыханова М.Г. (1981) 2020. Гнездование тростниковой суторы Paradoxornis heudei на восточном побережье озера Ханка в 1978 году // Рус. орнитол. журн. 29 (1974): 4297-4300. EDN: YIITEY.
• Назаров Ю.Н., Куринный В.Н. 1981. Новые встречи редких птиц в Приморском крае // Тр. Зоол. ин-та АН СССР 102: 110-111.
• Нечаев В.А., Гамова Т.В. 2009. Птицы Дальнего Востока России (аннотированный каталог). Владивосток: 1-564.
• Нечаев В.А., Горчаков Г.А. 2001. Гнездование тростниковой суторы Paradoxornis heudei David на побережье Японского моря // Животный и растительный мир Дальнего Востока. Уссурийск, 5: 151-155.
• Омелько М.А., Омелько М.М. Проблемы сохранения орнитофауны окрестностей г. Владивосток на примере полуострова Де-Фриза // 9-я Дальневост. конф. по заповедному делу. Владивосток: 312-317.
• Поливанов В.М., Поливанова Н.Н., Шибнев Ю.Б., Кудель А.Ф. 1973. Новый род и вид птицы в фауне СССР – тростниковый ополовник – Paradoxornis heudei David // Докл. АН СССР. Сер. биол. 212, 1: 257.
• Поливанова Н.Н., Поливанов В.М., Шибнев Ю.Б. 1974. О гнездовании тростникового ополовника (Paradoxornis heudei) на озере Ханка // Материалы 6-й Всесоюз. орнитол. конф. М., 2: 103-105.
• Поливанова Н.Н., Поливанов В.М., Шибнев Ю.Б. 1980. К биологии тростникового ополовника (Paradoxornis heudei polivanovi Stepanyan) // Бюл. МОИП. Отд. биол. 85, 6: 31-41.
• Поливанова Н.Н., Поливанов В.М., Шибнев Ю.Б. 2009. К биологии тростникового ополовника Paradoxornis heudei polivanovi // Рус. орнитол. журн. 18 (460): 119-131. EDN:
JVLLRZ.
• Романов А.Л., Романова А.И. 1959. Птичье яйцо. М.: 1-620.
• Сотников В.Н. 2023. Каталог коллекций. Птицы – Aves. Оологическая и нидологическая коллекции. Вып. 2. Воробьинообразные – Passeriformes. Киров: 1-304.
• Сотников В.Н., Ластухин А.А., Глущенко Ю.Н., Вялков А.В., Бачурин Г.Н., Мещерягина С.Г.,
Шибнев Ю.Б. 2016. Орнитологические наблюдения в Приморском крае в 2015 году // Рус. орнитол. журн. 25 (1269): 1151-1169. EDN: VOASBZ.
• Степанян Л.С. 1974. Paradoxornis heudei polivanovi Stepanyan ssp.n. (Paradoxornithidae, Aves) из бассейна озера Ханка // Зоол. журн. 53, 8: 1270-1272.
• Степанян Л.С. 1998. О видовой самостоятельности Paradoxornis polivanovi (Paradoxornithidae, Aves) // Зоол. журн. 77, 10: 1158-1161.
• Степанян Л.С. 2003. Конспект орнитологической фауны России и сопредельных территорий (в границах СССР как исторической области). М.: 1-808.
• Brazil M.А. 2009. Birds of East Asia. Eastern China, Taiwan, Korea, Japan and Eastern Russia.
London: 1-529.
• Del Hoyo J., Collar N.J. 2016. HBW and BirdLife International Illustrated Checklist of the Birds of the World. Vol. 2. Passerines. Barcelona: 1-1013.
• Gombobaatar S.B. 2009. A Dictionary of Vertebrate Animals of Mongolia. Ulaanbaatar: 1-318.
• Hoyt D.F. 1979. Practical methods of estimating volume and fresh weight of bird eggs // Auk 96:73-77.
• MacKinnon J., Phillipps K. 2000. A Field Guide the Birds of China. Oxford Univ. Press: 1-586.
• Robson C. 2007. Family Paradoxornithidae (Parrotbills) // Handbook of the Birds of the World. Vol. 12. Picathares to Tits and Chickadees. Barcelona: 292-320.
• The CNG Field Guide to the Birds of China. 2024. Hunan Science and Technology Press, Changsha: 1-686.
Юрий Николаевич Глущенко, Дмитрий Вячеславович Коробов. Тихоокеанский институт географии ДВО РАН, Владивосток, Россия. E-mail: yu.gluschenko@mail.ru, dv.korobov@mail.ru
Иван Михайлович Тиунов. ФНЦ биоразнообразия наземной биоты Восточной Азии ДВО РАН, Владивосток, Россия. Государственный природный биосферный заповедник «Ханкайский», Спасск-Дальний, Приморский край, Россия. E-mail: ovsianka11@yandex.ru
Владимир Несторович Сотников. Кировский городской зоологический музей, Киров, Россия. E-mail: sotnikovkgzm@gmail.com
Андрей Витальевич Вялков. Владивосток, Россия. E-mail: adrem-tan@yandex.ru
Источник
Русский орнитологический журнал 2024, экспресс-выпуск 2472. стр. 4771-4796.