

Корольковая пеночка Phylloscopus proregulus — обычный вид хвойной и смешанной тайги Сибири и Дальнего Востока. Тем не менее, некоторые стороны её биологии остаются почти не изученными. В полной мере это относится и к процессу смены оперения. Специальная работа имется только по послебрачной линьке корольковой пеночки из северных районов Буреинского хребта (Медведева 1999). В работах фаунистической направленности данные по линьке рассматриваемого вида носят отрывочный характер и не дают полного представления об этом явлении его годового цикла (Воробьёв 1963; Нечаев 1991). Более подробно в литературе освещены вопросы гнездовой биологии и сезонные перемещения Ph. proregulus (Воробьёв 1963; Панов 1973; Нечаев 1991; Бисеров 1999).
Буреинский хребет находится в северо-восточной части области гнездования корольковой пеночки. Мы изучали её в летне-осенний период в 3 равноудалённых друг от друга районах Буреинского хребта: 1) на северной оконечности хребта и сопредельных территориях — у слияния рек Левая и Правая Бурея (Буреинский заповедник, 550 м н.у.м., в 1997 году), верховья р. Ниман (правый приток Бурей, 1050 м н.у.м., в 1998); 2) в центральной части хребта, в среднем течении р. Дубликан (Дубликанский заказник, 300 м н.у.м., в 1999); 3) на южной оконечности хребта, в верховьях р. Икура (Заповедник “Бастак”, 150-200 м н.у.м., в 2000). Расстояние между крайними точками составило около 300 км.
Основным методом сбора материала служил отлов птиц паутинными сетями. Их общая протяжённость составляла 100 м. Сети стояли в пойменных лесах, существенно различающихся по видовому составу деревьев. В месте слияния Левой и Правой Бурей это были первичные смешанные (лиственница, пихта, ель, несколько видов тополей, ольха, берёза) и лиственные (тополёво-чозениевые) леса; в верховьях Нимана — первичные хвойно-лиственные (лиственница, пихта, ель, тополь, ольха, берёза) и заросли кедрового стланика с примесью жимолости; на Дубликане — вторичные лиственные (тополь, охьха, берёза, ива, черёмуха) леса с примесью лиственницы; на Икуре — вторичные лиственные леса (дуб, тополь, ива, берёза, липа).
Материал и методики
В работе использованы данные прижизненного обследования отловленных в паутинные сети корольковых пеночек. Помимо описания состояния перьевого покрова, у них оценивались величина подкожных жировых резервов, степень пневматизации черепа, развитие наседного пятна и клоакального выступа (у взрослых особей), измерялись длина крыла и, в случае дорастания рулевых у молодых, длина хвоста. Для выяснения возраста, в котором молодые птицы покидают гнёзда, а также состояния их оперения к моменту вылета, велись наблюдения за птенцами из 4 гнёзд с момента их вылупления.
Линьку описывали по методике, предложенной орнитологами Петербургского университета (Носков, Гагинская 1972; Гагинская, Рымкевич 1974; Носков, Рымкевич 1977). В основу выделения стадий послебрачной линьки положен метод, основанный на выделении 11 стадий, где первые 10 соответствуют смене первостепенных маховых, а 11-я — смене проксимальных второстепенных маховых и дорастанию контурного оперения. При выделении стадий постювенальной линьки использовали разницу в сроках начала и окончания смены пера на разных птерилиях. Использовали сквозную нумерацию маховых, считая от дистального пера. Полноту смены оперения на отдельных птерилиях и их участках характеризовали соответствующим показателем (Рымкевич и др. 1987).
Для оценки индивидуальной продолжительности послебрачной и постювенальной линек использовали как прямые наблюдения в природе за окольцованными птицами, так и косвенные показатели (промежуток времени между отловами первой линяющей и первой перелинявшей особи). Под сезоном линьки мы понимаем период, в течение которого в исследуемом районе в каждый конкретный год встречаются линяющие корольковые пеночки. При обобщении данных за несколько лет и из разных точек Буреинского хребта сезон линьки именовался суммарным. О начале осеннего пролёта и его динамике судили по числу отлавливаемых ежедневно особей.
По нашим данным, у молодых корольковых пеночек пневматизация черепа не завершается к концу осенней миграции. Это позволяет безошибочно отличать их от особей старше года на протяжении всего периода их пребывания в районе наших исследований.
Всего за период исследований осмотрены 304 корольковые пеночки: 73 взрослых (37 в состоянии линьки) и 241 молодая (182 линяющих). Большая часть данных по линьке и взрослых, и молодых птиц собрана в северной части Буреинского хребта, где корольковая пеночка входит в число доминирующих видов и во множестве гнездится в пойменных чозениево-тополёвых лесах и зарослях кедрового стланика.
Послебрачная линька
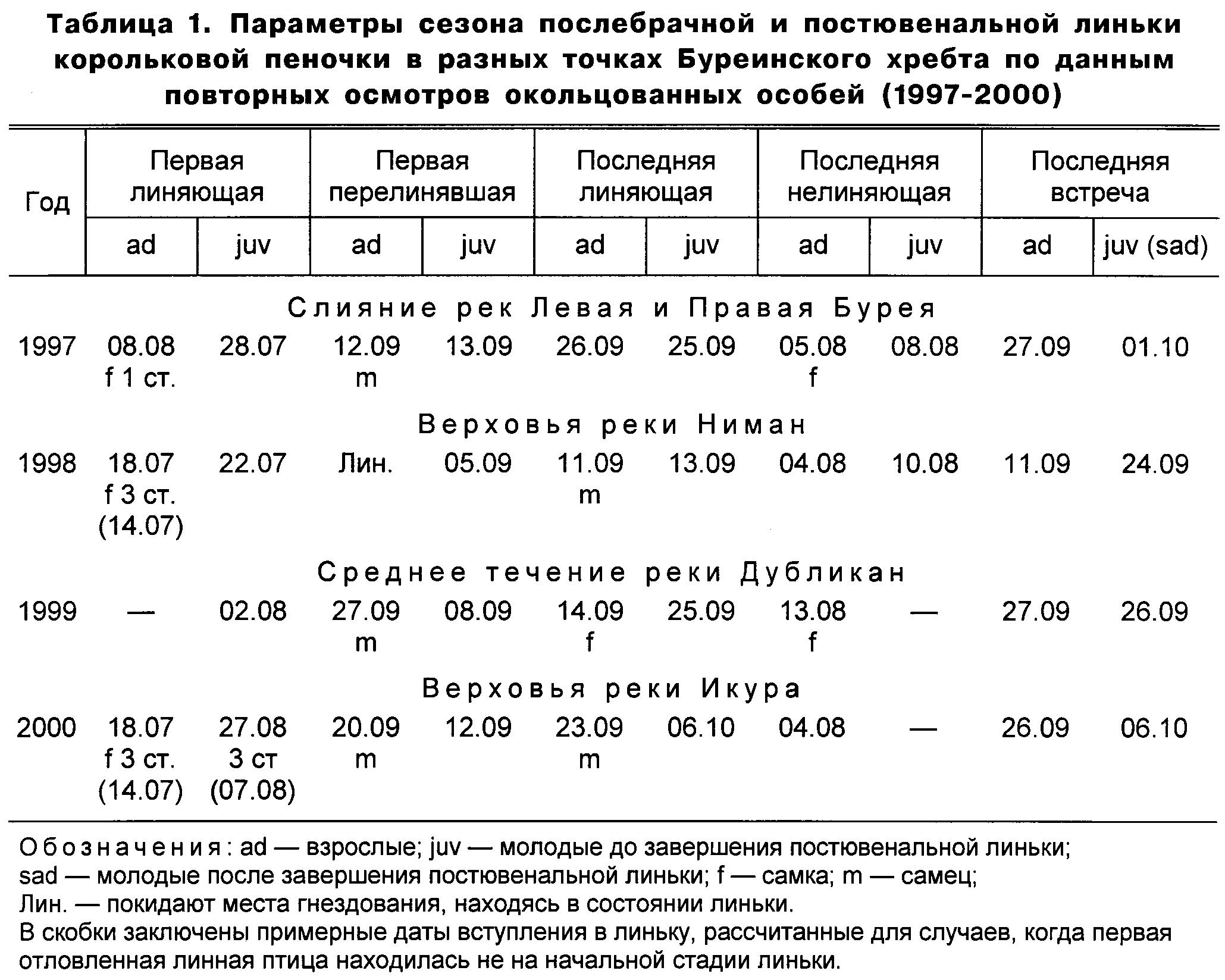
О сроках и продолжительности сезонов послебрачной линьки корольковой пеночки в разных точках Буреинского хребта можно судить по данным таблицы 1. Суммарный сезон послебрачной линьки на Буреинском хребте составляет 75 дней и длится с 14 июля по 26 сентября. Сроки прохождения корольковыми пеночками буреинской популяции раных стадий послебрачной линьки показаны на рисунке 1.
Первыми начинают линять самки, успешно завершившие гнездовой цикл (на Буреинском хребте у этого вида одна кладка в сезоне) или утратившие потомство на стадии птенцов и слётков. Среди последних ещё не линяющих птиц в отловах также были одни самки. Такая растянутость времени вступления в линьку у самок корольковых пеночек (не менее 20 дней) объясняется повторным размножением после гибели первой кладки.
В отличие от самок, самцы корольковой пеночки вступают в линьку почти синхронно в конце второй-начале третьей декады июля, что обусловлено тем, что они не участвуют в выкармливании птенцов.
Самки корольковой пеночки не совмещают послебрачную линьку с инкубацией, кормлением птенцов и слётков младше 18-20 сут. Так, три самки, отловленные с 13-18-сут слётками (длина растущих рулевых перьев последних 20-28 мм) 19 июля 1998, 4 августа 1998 и 5 августа 1997, к линьке ещё не приступили. Самка, отловленная 28 июля 1998 с 20-22-сут слётками (длина рулей 33-35 мм), находилась на 2-й стадии послебрачной линьки, причём линька у неё началась не более 2 дней назад (9-е и 10-е маховые были представлены одинаковыми еле заметными трубочками). Самка, у которой 25 июля было разорено гнездо с птенцами, 27 июля была отловлена с выпавшими десятыми маховыми перьями.
Самки покидают гнездовые участки вместе с выводками в течение недели после вылета птенцов из гнезда и начинают линьку, как правило, за пределами гнездовых участков.
Большая часть самцов линяет на своих гнездовых участках, переставая в этот период петь. Пение вновь возобновляется в конце августа-начале сентября, когда послебрачная линька самцов достигает 9-й или 10-й стадии. Гнездовые участки самцы покидают в середине первой-начале второй декады сентября, находясь на 10-11 стадиях послебрачной линьки.
У корольковой пеночки послебрачная линька полная, показатель полноты смены оперения на всех птерилиях равен 1. Порядок вступления птерилий и их участков в линьку показан в таблице 2. Индивидуальная продолжительность послебрачной линьки, по данным повторных отловов, составила 55-60 дней (рис. 1).
Постювенальная линька
Наиболее ранняя дата отлова только что покинувших гнездо слётков — 4 июля (1997), но основная масса таких встреч приходилась на период с 15 июля по 4 августа. Птенцы покидают гнёзда в возрасте 13-15 сут. В это время у них наблюдается интенсивное дорастание основной и некоторых групп дополнительной частей юношеского оперения: маховых, рулевых, перьев туловищных птерилий, нижних кроющих крыла.
Линять молодые начинают в возрасте 25-30 сут, причём примерно у 50 % осмотренных нами особей к этому времени ещё не завершился рост рулевых


перьев (длина 36-40 мм). Незадолго до начала линьки или одновременно с ней начинается рост дополнительной части юношеского оперения: периферийных перьев туловищных птерилий, голени, перьев бокового отдела, а также нижних плечевых и маргинальных, пекторальных, части верхних и нижних кроющих пропатагиальной складки, наружного ряда клоакальных перьев, пуховидных перьев на юношеских аптериях.
Постювенальная линька корольковой пеночки частичная. Её удаётся разделить на 5 стадий:
1) линька только на туловищных птерилиях;
2) начинает линять лобно-затылочный отдел головной птерилии и голень, идёт зарастание пуховидными перьями юношеских аптерий;
3) разгар линьки лобно-затылочного отдела головной птерилии (ювенильные перья отсутствуют) и голени, зарастание аптерий завершено;
4) продолжается линька лобно-затылочного отдела, на голени линька завершена;
5) закончена линька лобно-затылочного отдела, завершается линька на других птерилиях (дорастающие перья имеют вид больших кисточек).
Порядок вступления в линьку и полнота замены перьев на них показаны в таблице 3. Как видно, наименее подвержена линьке крыловая птерилия, где полностью заменяются только нижние плечевые и пекторальные перья.

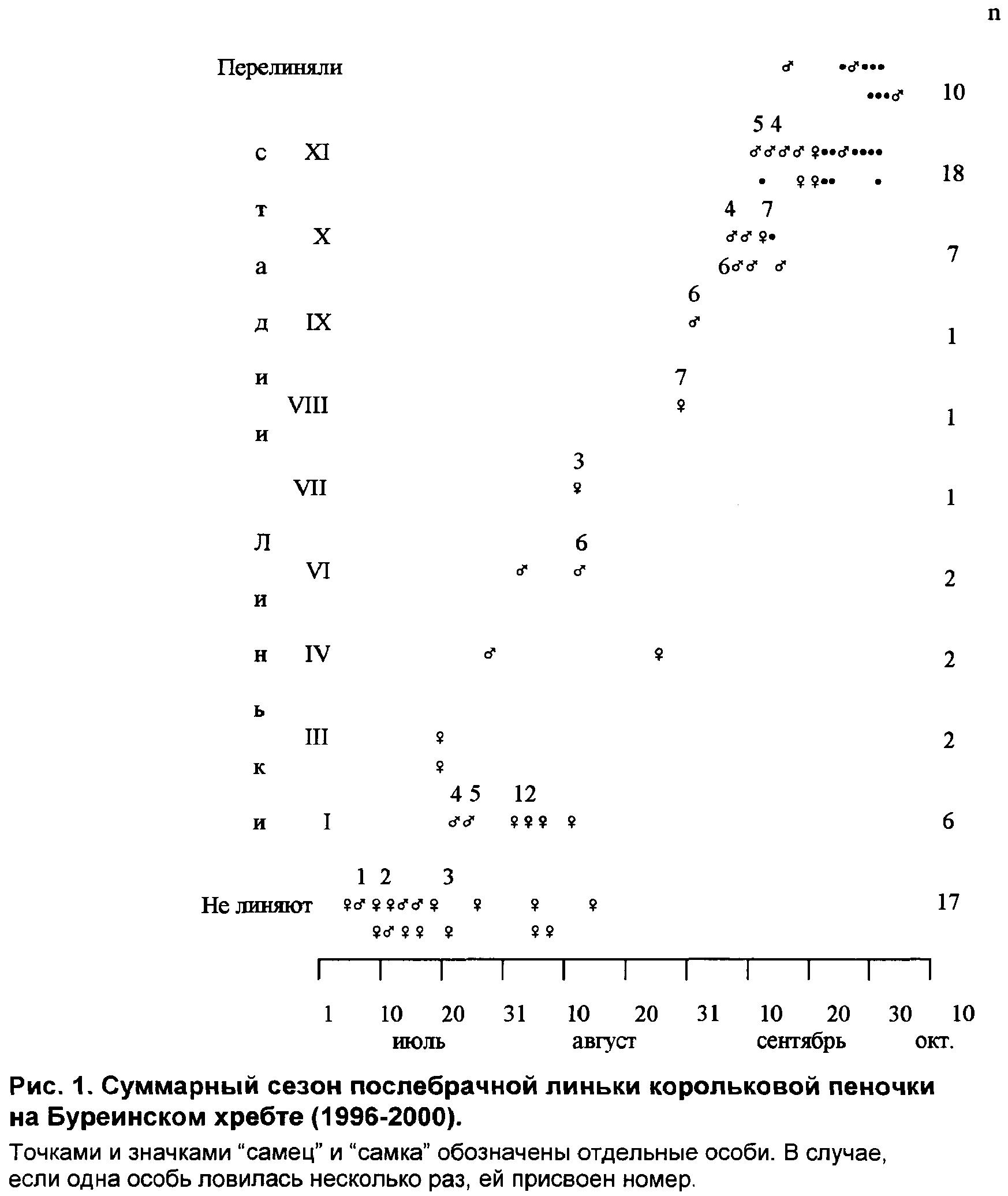
Основные характеристики сезонов постювенальной линьки приведены в таблице 1. Период встречаемости линных особей в разных точках Буреинского хребта составил 41-60 дней, а по хребту в целом — около 77 дней.
Молодые корольковые пеночки покидают гнездовые участки ещё находясь в выводках. В дальнейшем они не оседают надолго на одном месте, даже на время линьки, что подтверждается отсутствием повторных отловов. За неимением последних о продолжительности постювенальной линьки мы судили только по косвенным данным, согласно которым индивидуальная продолжительность постювенальной линьки оценена в 37-48 дней (рис. 2).
Осенняя миграция
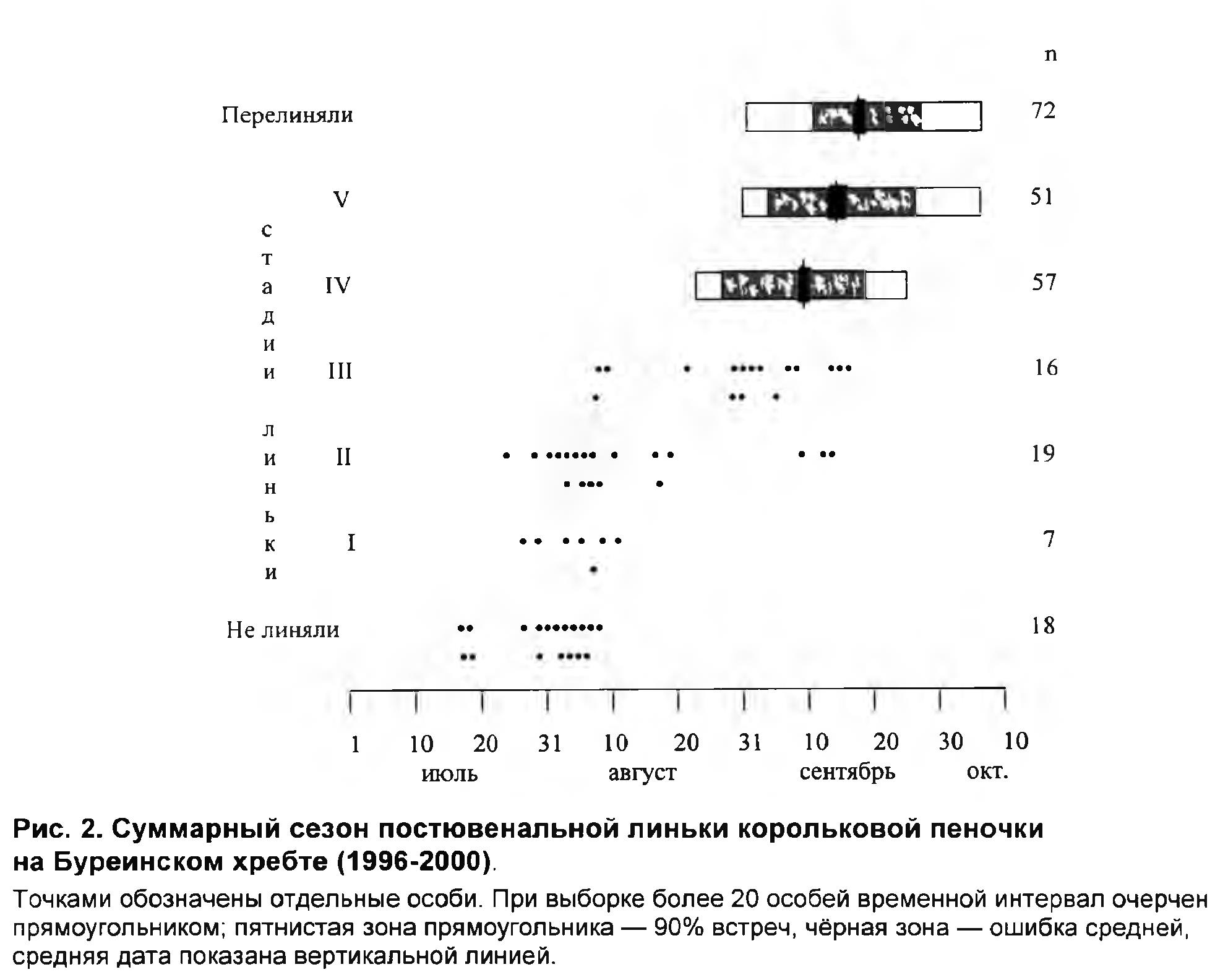
Начало осеннего пролёта корольковой пеночки на Буреинском хребте в 1997-2000 приходилось на разные числа первой половины сентября (рис. 3). В верховьях Нимана (1050 м н.у.м.; территория, примыкающая к северным районам Буреинского хребта) осенний пролёт был выражен слабо и скорее походил на отлёт птиц местной популяции. Пролёт в центральной части хребта был сходен по срокам и динамике с пролётом в северной его части на высоте 550 м н.у.м., но был более массовым. В южной части хребта все сроки осенней миграции (начало, пик, окончание) запаздывали на 5-7 дней, по сравнению с центральной и северной частью. Во все годы наблюдений осенний пролёт корольковых пеночек проходил в виде одной волны с пиком в третьей (1997, 1999) или четвёртой (2000) пятидневках сентября.
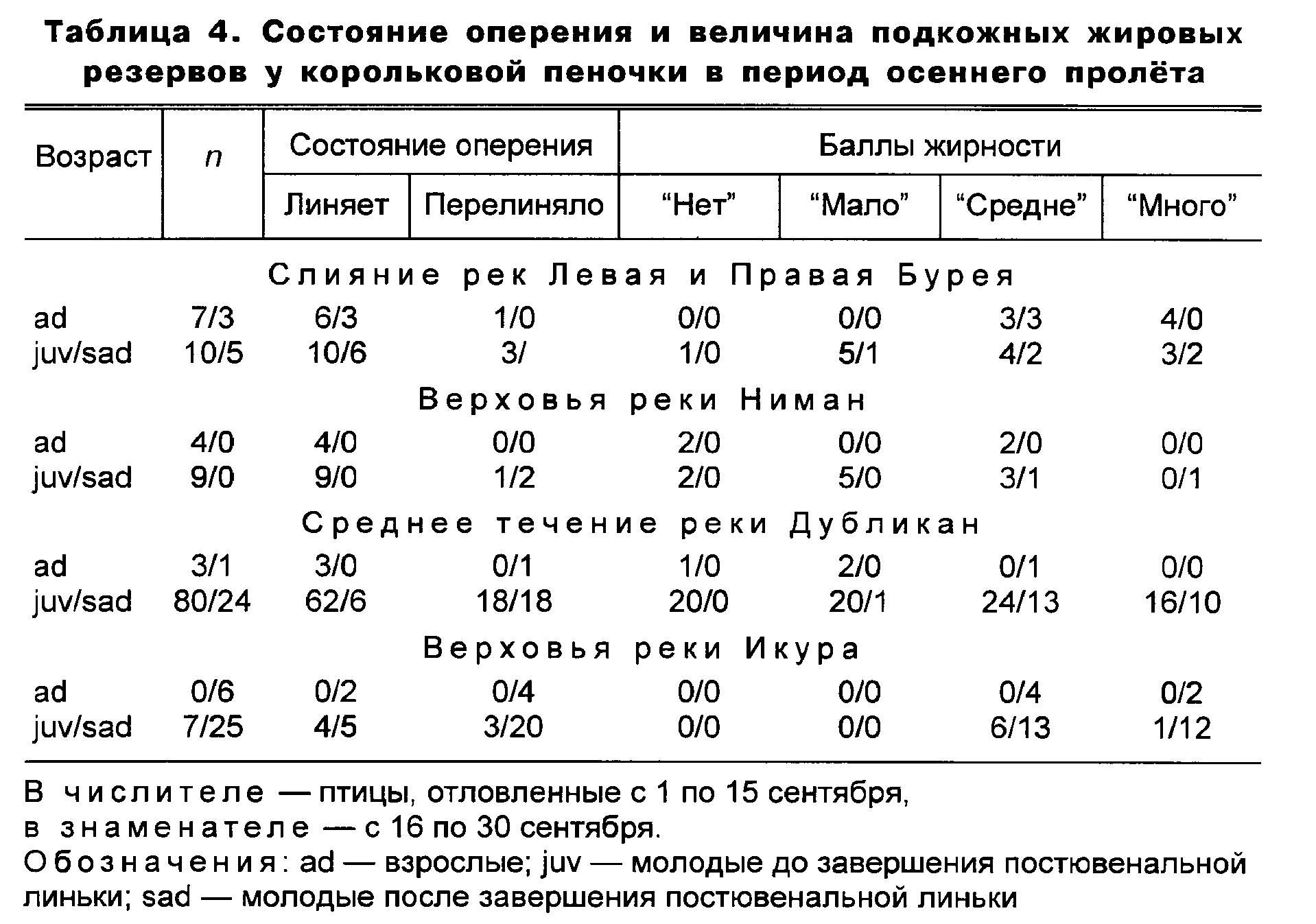
Значительная часть как взрослых, так и молодых корольковых пеночек совмещала осеннюю миграцию с завершающей стадией линьки (табл. 4). Особенно много таких особей было в первую половину пролёта. В первой половине сентября доля пролётных пеночек, завершающих постювенальную линьку, составляла 57-90 %, во второй половине сентября этот показатель снижался до 0-60 %. Соответственно, возрастала доля птиц, завершивших линьку.
Если обобщить данные по состоянию запасов в подкожных жировых депо у мигрирующих молодых корольковых пеночек по всему Буреинскому хребту (табл. 4), то видно, что в первую половину сентября количество особей с баллами жирности “нет”-“мало” и “средне’’-“много” примерно одинаково (табл. 4). Во второй половине сентября явно доминировали средне и максимально жирные птицы (91 %).
Литература
• Бисеров М.Ф. 1999. О гнездостроении корольковой пеночки на северо-востоке ареала // Тр. заповедника “Буреинский” 1: 63-67.
• Воробьёв К.А. 1963. Птицы Якутии. М.: 1-336.
• Гагинская А.Р., Рымкевич Т.А. 1976. Методические указания к прижизненной обработке птиц // Материалы 5-го заседания рабочей группы по проблеме «Исследование продуктивности вида в пределах ареала». Вильнюс: 155-166.
• Медведева Е.А. 1999. Послебрачная линька корольковой пеночки // Тез. докл. 4-й Дальневосточной конф. по заповедному делу. Владивосток: 104-105.
• Нечаев В.А. 1991. Птицы острова Сахалин. Владивосток: 1-762.
• Носков Г.А., Рымкевич Т.А. 1977. Методика изучения внутривидовой изменчивости линьки у птиц // Методики исследования продуктивности и структуры видов птиц в пределах их ареалов. Вильнюс: 37-48.
• Панов Е.Н. 1973. Птицы Южного Приморья. Новосибирск: 1-376.
• Рымкевич Т.А., Могильнер А.И., Носков Г.А., Яковлева Г.А. 1987. Новые показатели для характеристики линьки воробьиных птиц // Зоол. журн. 66, 3: 444-453.
Источник
Русский орнитологический журнал 2002, Экспресс-выпуск 188. стр. 551-560.