

Федеральное государственное бюджетное образовательное учреждение высшего образования
Московский государственный университет имени М.В. Ломоносова
Биологический факультет
На правах рукописи
Антипов Владислав Анатольевич
Диссертация на соискание ученой степени кандидата биологических наук
Научный руководитель доктор биологических наук профессор В.В. Иваницкий
Москва 2016
Содержание (часть 2)
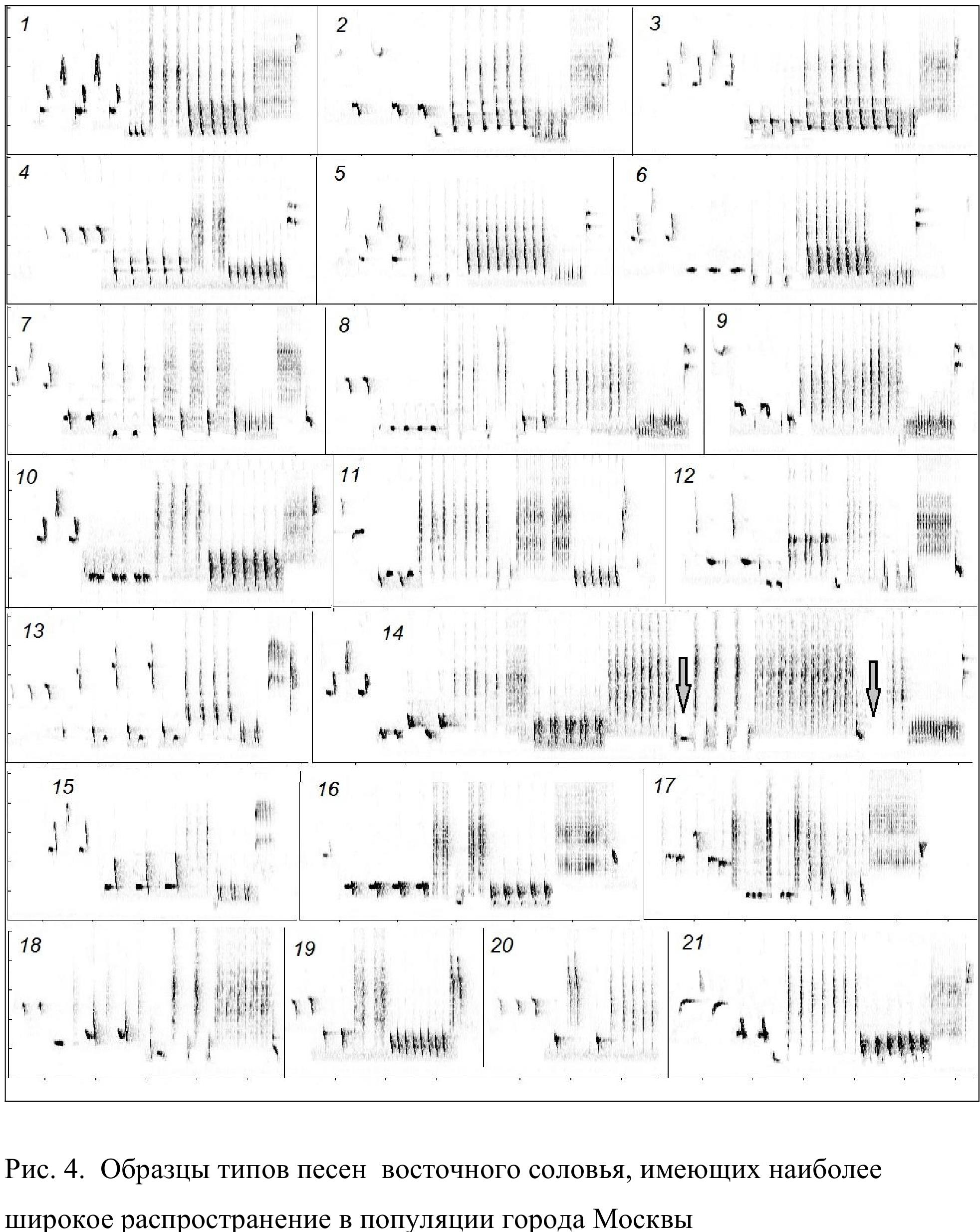
Глава 3. Фонетические особенности рекламной вокализации, структура отдельных песен и объем индивидуальных репертуаров восточного соловья
3.1. Принципы описания песни и терминологии
3.2. Фонетические особенности и структура единичных песен
3.3. Количественные параметры единичных песен (строф)
3.4. Объем индивидуальных репертуаров и локальное разнообразие типов песен
Глава 4. Синтаксическая организация в песне восточного соловья: очередность исполнения и группирование типов песен
4.1. Структура и сходство индивидуальных репертуаров
4.2. Совместная встречаемость разных типов песен в репертуарах
4.3. Переключения между типами песен по ходу вокальных сессий
4.4. Секвенциональная организация вокальных сессий
4.5. Поведенческие особенности исполнения программ
Глава 3. ФОНЕТИЧЕСКИЕ ОСОБЕННОСТИ РЕКЛАМНОЙ ВОКАЛИЗАЦИИ, СТРУКТУРА ОТДЕЛЬНЫХ ПЕСЕН И ОБЪЕМ ИНДИВИДУАЛЬНЫХ РЕПЕРТУАРОВ ВОСТОЧНОГО СОЛОВЬЯ
3.1 Принципы описания песни и терминология
Как и у большинства других певчих птиц, рекламная вокализация восточного соловья может быть описана как многоуровневая система с иерархической организацией. В роли элементарных акустических единиц выступают ноты (элементы); на сонограмме при любом разрешении они выглядят, как монолитные, далее не делимые конструкции, не содержащие внутри себя пауз.
Приемы группирования нот на следующем уровне организации песни варьируют. Часто разные ноты попарно объединяются в слоги (фигуры) – устойчивые конструкции, которые всегда исполняются как целое. Порознь такие ноты обычно никогда не исполняются. Промежуток между элементами, составляющими слог, иногда составляет всего 15 – 20 мс, и заметить его можно лишь при большом разрешении (т.е. при «растягивании» сонограммы по оси времени). Если же группируются друг с другом несколько одинаковых нот или одинаковых слогов, то образуются фразы (секции). В одной песне может быть от 3 до 8 фраз (медиана 5). Наиболее распространенным типом фраз у восточного соловья являются разнообразные трели. В отличие от фигур число нот (слогов) во фразах обычно широко варьирует от случая к случаю. Одиночные ноты, не входящие в состав слогов или фраз, встречаются редко, но для некоторых вариантов песни они весьма характерны.
На следующем уровне интеграции находятся отдельные (единичные) песни. Каждая единичная песня представляет собой дискретную и стереотипную вокальную конструкцию, четко ограниченную паузами, состоящую из фраз (секций), слогов, отдельных нот и относящуюся к определенному типу песен. Последовательность разнотипных песен, исполняемых данным самцов подряд и записанных на фонограмме, мы будем называть вокальной сессией (песенным циклом). Очередность исполнения разных типов песен по ходу вокальной сессии мы будем называть синтаксической организацией; типы песен, обычно исполняемые друг за другом в определенной последовательности – синтаксически связанными типами песен, а повторяющиеся (стереотипные) последовательности исполнения синтаксически связанных типов песен – программами (Иваницкий и др., 2013; Антипов, 2011, 2012, 2013).
3.2 Фонетические особенности и структура единичных песен
Одна из наиболее характерных фонетических особенностей песни восточного соловья состоит в том, что она занимает очень широкий частотный диапазон – от 1,2 до 10 кГц (рис. 3). В основном песня состоит из широкополосных и очень коротких нот и слогов длительностью всего 10 – 30 мс, занимающих почти весь вышеуказанный диапазон. Но кроме них, песня соловья включает в себя и относительно низкочастотные трели, локализованные в более узкой полосе – от 1 до 2-3 кГц (рис. 3Д). В этом же диапазоне локализованы и некоторые одиночные ноты, придающие своеобразный ритмический рисунок некоторым типам песен (рис. 3В). Такие низкочастотные трели и одиночные ноты представляют собой тоновые сигналы.
Интересно, что пение соловья насыщено звуками, лежащими ниже 2 – 2.5 кГц. Такие низкочастотные звуки редко присутствуют в пении мелких певчих птиц. Крайне медленно затухая, они распространяются на большое расстояние (сотни метров) и придают соловьиному пению специфическое звучание. В его основе лежит акустический контраст между фразами, составленными из трелей разного частотного диапазона. При этом птица очень быстро переключается между этими сильно отличающимися вокальными компонентами.

Единичные песни восточного соловья довольно продолжительны (до 10 – 14 с) и имеют своеобразную, подчас сложную структуру. Некоторые из них включают до 40 – 50 разнообразных компонентов (нот и слогов), сгруппированных в несколько (до 5 – 6) фраз. Почти перед каждым типом песен восточного соловья присутствует серия специфических сигналов. Их принято называть «почин» (рис. 3А). «Почины» локализованы в диапазоне от 4 до 8 кГц. Они составляют самую высокочастотную часть соловьиного пения и слышны лишь на близком расстоянии. Это тоновые сигналы, иногда двусложные (состоят из двух нот) и разнообразные по характеру частотной модуляции. Перед одной песней подряд могут быть исполнены до 8-10 одинаковых «починов», так что иногда они практически целиком заполняют все промежутки между песнями. Но если промежутки сокращаются до минимума, «почины» не используются вовсе. В московской популяции, изученной наиболее подробно, отмечены 7 типов «починов». То есть, их разнообразие значительно уступает разнообразию типов песен. По ходу вокальной сессии одинаковые «почины» обычно исполняются подряд перед несколькими (обычно перед 3 – 4) разными типами песен, следующими друг за другом. Таким образом, в исполнении починов отчетливо выражена тенденция к периодической вариативности, тогда как сами песни по ходу вокализации сменяются практически непрерывно. Не исключено, что за генерацию починов и песен ответственны разные нейронные механизмы. Мы полагаем, что особенности исполнения «починов» свидетельствуют о том, что они не являются составной частью типов песен. Вероятно, «почины» – это особый вокальный феномен, аналоги которого мы встречаем и в пении некоторых других видов, склонных заполнять промежутки между песнями равномерно повторяющимися, короткими и относительно тихими позывками. Хорошо известным примером может служить пение пеночки-теньковки (Phylloscopus collybita) (Симкин, 1990).
Собственно песня восточного соловья начинается, как мы полагаем, с «запевки» – серии из громких и мелодичных2 свистовых нот, лежащих в довольно узком и низком частотном диапазоне 1 – 3 кГц (рис. 3Б). По звучанию «запевки» обычно (хотя и не всегда) контрастируют с тихими высокочастотными «починами» и со следующей – центральной частью песни, состоящей в основном из широкополосных нот. Но главное различие состоит в том, что запевка всегда специфична и исполняется лишь в составе данного типа песен, тогда как один и тот же почин исполняется перед
разными типами песен, следующими друг за другом. То есть, почины обладают некоторой инерционностью.
В центральной части песни преобладают трели – серии коротких (обычно широкополосных) звуков разной природы. Они могут группироваться в разреженные серии. Мы воспринимаем их на слух как трещание. Многие звуки, образующие трели, представляют собой слоги, то есть состоят из двух посылок (нот), разделенных очень коротким интервалом около 15 – 30 мс. Первой всегда исполняется низкоамплитудная шумовая нота. За ней следует значительно более громкая тоновая нота в диапазоне 1.2 – 3.0 кГц. Общая продолжительность этой двусложной конструкции около 100 мс, длина разделяющей паузы около 30 мс (рис. 3Г).
Финал песни обычно содержит две четко обособленные части. Вопервых, это специфическая трель – очень плотная (50-55 в секунду) упаковка («пачка») широкополосных импульсов общей продолжительностью до 0.5 с (негромкое жужжание или скрипение) (рис. 3Е). Такие «пачки» всегда исполняются только в конце песни. Во-вторых, это очень короткий, но четко выраженный замыкающий компонент, маркирующий конец песни (рис. 3Ж). Обычно он исполняется сразу же после вышеупомянутой трели. Продолжительность таких компонентов около 100 мс. Некоторые из них представляют собой одиночную тоновую, частотно модулированную ноту. Другие включают две такие ноты, исполняемые одновременно в режиме бифонации. Всего в изученной нами популяции отмечено 6 разных типов замыкающих компонентов. Преобладают высокочастотные сигналы в диапазоне 5-8 кГц, но отмечен и один низкочастотный вариант (1.5–4.5 кГц).
Таким образом, многие типы песен восточного соловья строятся по единому плану. Обычно песня предваряется «почином» и начинается с «запевки» – композиции из звучных низкочастотных свистовых нот. Затем исполняется основная часть песни, построенная преимущественно из широкополосных элементов, сгруппированных в плотные серии одинаковых нот. Завершают песню финальная трель и замыкающий компонент.
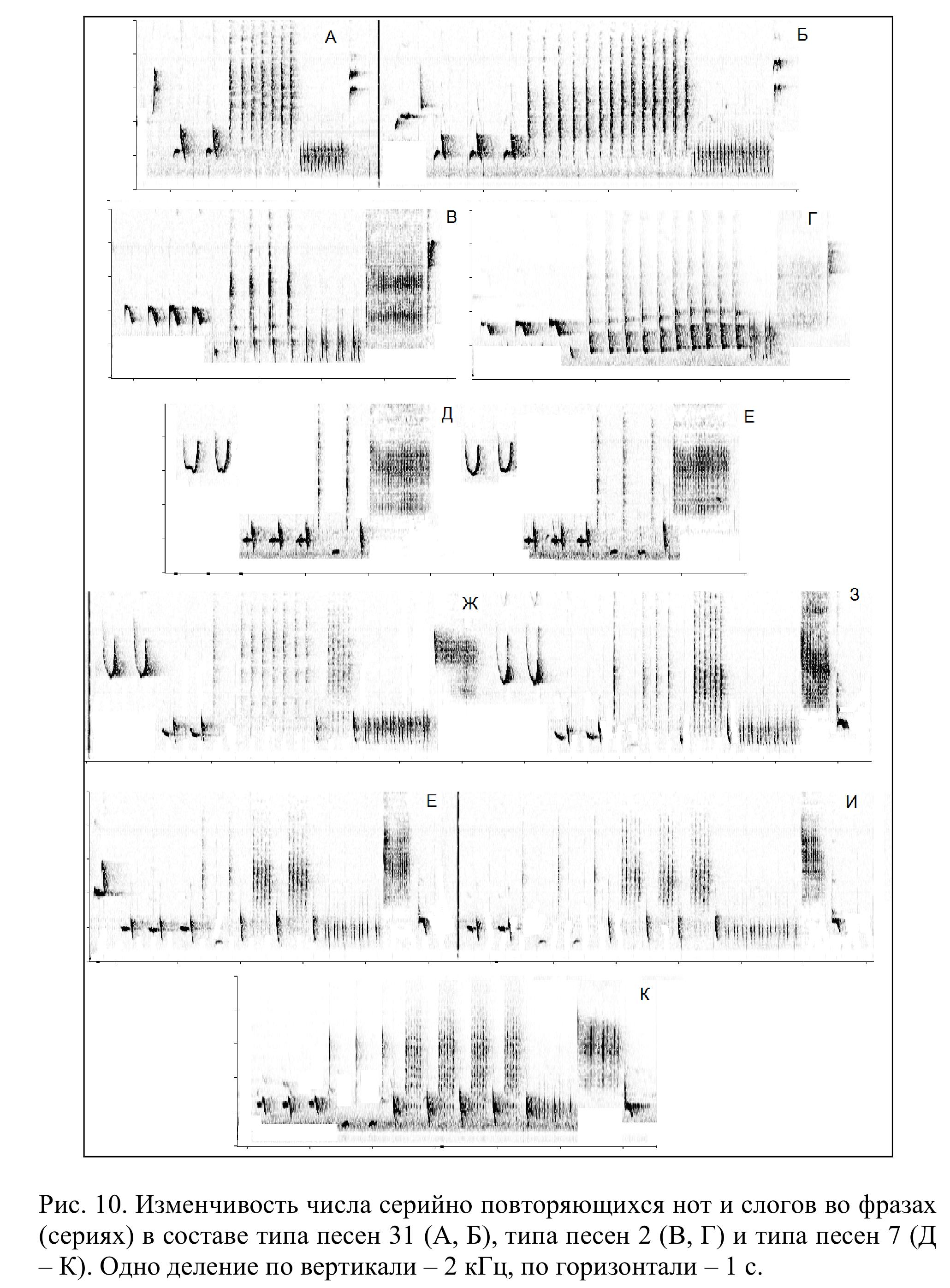
Как показано на рис. 4 и 10, в структуре некоторых типов песен допускаются отступления от описанного плана. Возможна, например, редукция центральной части (типы песен 4, 13, 15) или, напротив, ее существенное усложнение (14), а также разделение на две и более части за счет внедрения низкочастотных компонентов (11). Некоторые самцы склонны к усложнению начальных и финальных отделов песни за счет добавления дополнительных низкочастотных компонентов (типы песен 2, 3, 12, 13, 21). Таким образом, упомянутое выше деление песни восточного соловья на отдельные части (начальную, среднюю, заключительную) в целом весьма условно.
Как уже отмечалось, многие песни соловьи исполняют в усеченном виде. Важно, что при этом всегда исполняются лишь самое начало песни. Чаще всего это «запевка», присутствующая, например, в типах песен 1, 10, 14, 15, 16 и 17, иногда вместе с несколькими широкополосными звуками из центральной части песни. Автономного исполнения прочих компонентов песен (трелей всех типов и замыкающих элементов) не отмечено ни разу. То есть, соловей всегда берется за исполнение очередной песни с самого ее начала. Нередко несколько усеченных одинаковых или разных песен исполняются подряд.
3.3 Количественные параметры единичных песен (строф)
Для измерения продолжительности отдельных песен и пауз между ними, мы сформировали выборку из 400 песен и следующих за ними пауз. Чтобы нивелировать возможные индивидуальные и локальные различия, мы включили в выборку по 8 измерений от 10 самцов из 5 локалитетов (Москва и самые удаленные от нее участки. Измеряли только полные (завершенные) и только разные песни, выбранные из фонограммы в случайном порядке. Длину песни измеряли без починов, от начала первой запевки до замыкающего компонента. Паузу измеряли от конца замыкающего компонента до первого почина перед следующей песней.
Полученные данные представлены на pис. 5. Продолжительность единичных песен варьирует в широких пределах: от 1.9 до 14.1 (в среднем 5.8 ± 2.1) с; продолжительность пауз между песнями – от 0.15 до 7.3 (в среднем 2.6 ± 1.0) с. Таким образом, паузы намного короче песен. Иногда песни даже следуют одна за другой почти без перерыва (Симкин, 1981).
На рис. 9 приведены сведения о продолжительности пауз между отдельными песнями 4-х самцов, певших с наибольшей скоростью, в режиме «слитной» вокализации. Как можно видеть, большая часть таких пауз лежит в том же самом интервале (от 100 до 300 мс), что и промежутки между вокальными компонентами внутри песни (рис. 5). Таким образом, при «слитном» пении паузы между песнями и паузы между вокальными компонентами внутри этих песен становятся почти одинаковыми. Пение становится практически сплошным, причем подобный ритм исполнения песни соловей поддерживает достаточно долго (десятки минут). Но, как правило, песни при любом темпе их исполнения сохраняют типичную структуру, и на сонограмме их разграничение обычно не вызывает затруднений. Таким образом, даже при самом интенсивном пении восточный соловей исполняет отдельные песни от начала и до конца, хотя заполнение окального ряда нередко отличается очень высокой плотностью, и промежутки между песнями сокращаются до минимума.


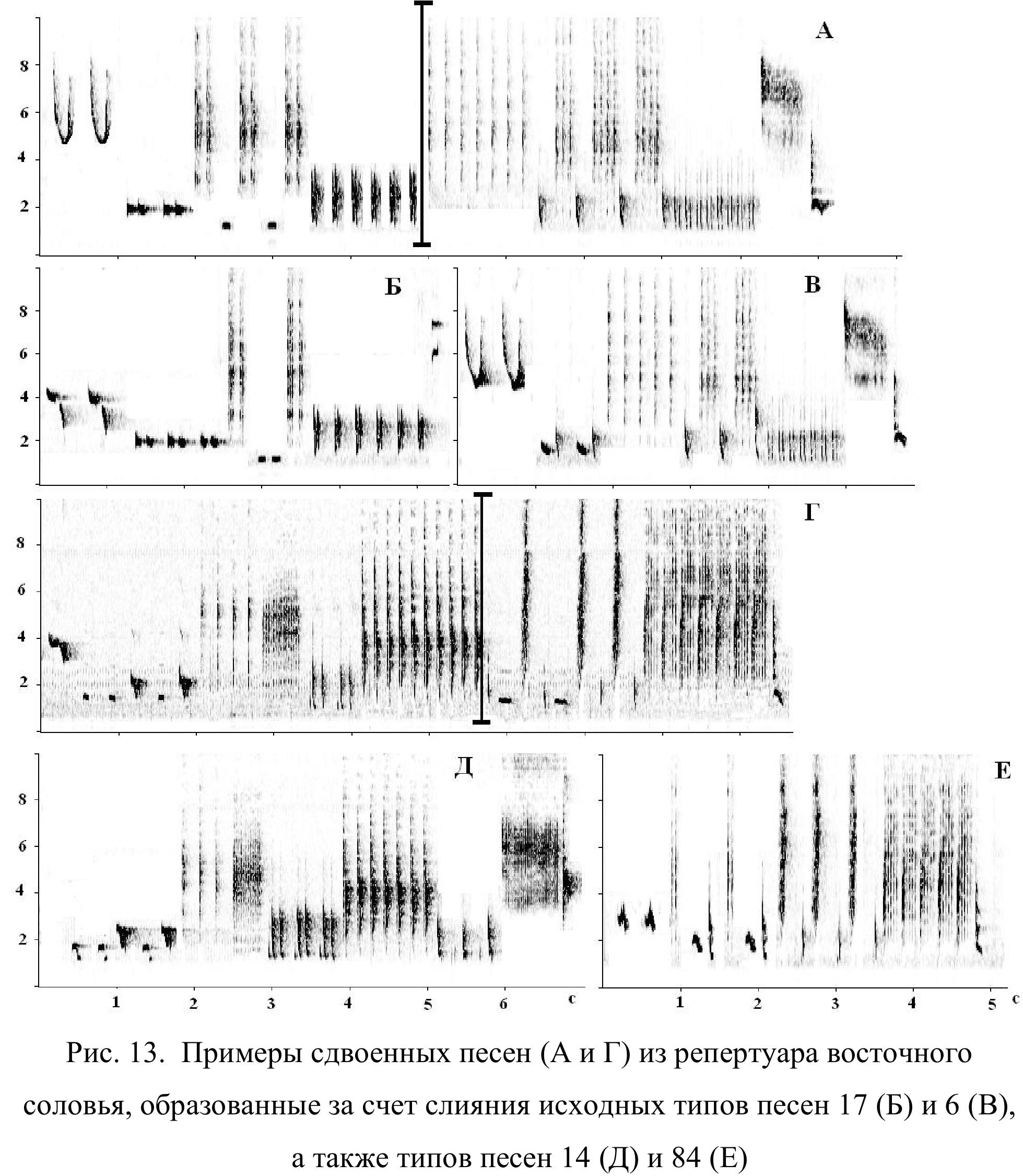
Что касается настоящего объединения (слияния) разнотипных песен, то в репертуаре восточного соловья оно не представляет редкости и, повидимому, составляет распространенный механизм образования новых типов песен. Причем тенденция к объединению песен не связана с темпом пения и, обычно, проявляется как индивидуальная особенность исполнителя. Пример такой конструкции (конгломерата) изображен на рис. 6. Обычно объединяются две синтаксически связанные песни, которые всегда исполняются друг за другом в определенном порядке (например, А → Б). В этом же порядке происходит и объединение. При этом первая песня (песня А) утрачивает свои финальные компоненты, а вторая песня (песня Б) лишается почина, а иногда и запевки. Чаще всего сдваивание песен носит факультативный характер. То есть, объединенная песня исполняется по ходу вокальной сессии наряду с исходными типами песен.

Данные о продолжительности вокальных компонентов, составляющих песню (нот, слогов и фраз), и пауз между ними приведены на рис. 7. По результатам 350 измерений на фонограммах 10 самцов продолжительность нот и слогов варьирует от 70 до 600 мс (в среднем 340 ± 120 мс), а продолжительность пауз – от 30 до 260 мс (в среднем 130 ± 50 мс). Эти два параметра связаны достоверной положительной корреляцией (рис. 8).


За более протяженными нотами (слогами) обычно следуют и более продолжительные паузы (Rsp = 0,56; p <0,001). В состав одной фразы входит до 22 нот или слогов (в среднем 4,7 ± 3,8; n = 400). Продолжительность фраз меняется от 60 до 2400 мс (в среднем 0,98 ± 0,51 мс; n = 400), продолжительность пауз, разделяющих фразы, от 50 до 500 мс (170 ± 120 мс). Таким образом, внутри песни (строфы) паузы между фразами несколько длиннее, чем паузы между нотами и слогами (t-test; p <0,001).
3.4 Объем индивидуальных репертуаров и локальное разнообразие типов песен
Объемы 388 индивидуальных репертуаров восточных соловьев варьируют от 2 до 21 типов песен (медиана – 9, нижний квартиль – 8, верхний квартиль – 11) (рис. 14). При этом 95 % всех репертуаров включают от 5 до 16 типов песен, и почти половина (51 %) их содержат 8, 9 или 10 типов песни. Таким образом, хотя общий размах изменчивости по этому параметру выглядит довольно широким, большинство исследованных нами птиц имеют сходный объем репертуара.
На рис. 15 приведены данные о репертуарах соловьев из разных участков обследованной нами территории. Согласно непараметрическому тесту Крускал-Уоллиса, влияние фактора принадлежности к участку достоверно (P < 0,01). Но вместе с тем, число достоверных парных различий (при введении поправки Ньюмена-Келси или поправки Бонферрони против множественных сравнений) очень невелико. Несколько выделяется в этом отношении лишь участок 9 (восточные районы Москвы). В то же время, репертуары московских городских соловьев, взятые в целом, больше, чем в области, и эти различия высоко достоверны (см. рис. 16) (тест Манн-Уитни, P < 0,001).
Поющий соловей представляет свой репертуар быстро. Этому способствует непрерывная вариативность пения, когда каждая очередная песня отличается от только что исполненных песен. Рис. 17 иллюстрирует ввод новых типов песен по ходу вокальных сессий 12 самцов. Данные свидетельствуют о том, что для относительно полного выявления состава индивидуального репертуара восточного соловья достаточно 40 – 50 песен. После этого линия аккумуляции выходит на плато.







Также интерес представляет скорость аккумуляции новых типов песен по мере увеличения числа анализируемых репертуаров самцов, обитающих на данной территории. Сколько соловьев на ней требуется записать для того, чтобы считать разнообразие типов песен выявленным достаточно полно ? Мы составили кривые аккумуляции для 5 участков, где было записано больше 25 самцов. Результаты представлены на рис. 18. Накопление новых типов песен вначале идет быстро. Суммарные репертуары для первых 10 самцов на разных участках содержали от 16 до 26 типов песен (в среднем для всех участков 21 тип песен). Дальнейшее увеличение числа репертуаров прибавляет относительно немного, а после 14 – 15 самцов кривая аккумуляции приближается к горизонтали. Таким образом, 10 – 15 репертуаров достаточно для относительно полного выявления разнообразия типов песен на данной территории. Отметим, что за единственным исключением (в секторе 2 было записано 11 соловьев) число самцов, записанных в одном секторе, было ≥ 20 (в среднем 32,3 ± 16,4). Это дает основания надеяться, что совокупное разнообразие типов песен на всех участках было выявлено нами с достаточной полнотой.
ГЛАВА 4. СИНТАКСИЧЕСКАЯ ОРГАНИЗАЦИЯ В ПЕНИИ ВОСТОЧНОГО СОЛОВЬЯ: ОЧЕРЕДНОСТЬ ИСПОЛНЕНИЯ И ГРУППИРОВАНИЕ ТИПОВ ПЕСЕН
4.1. Структура и сходство индивидуальных репертуаров
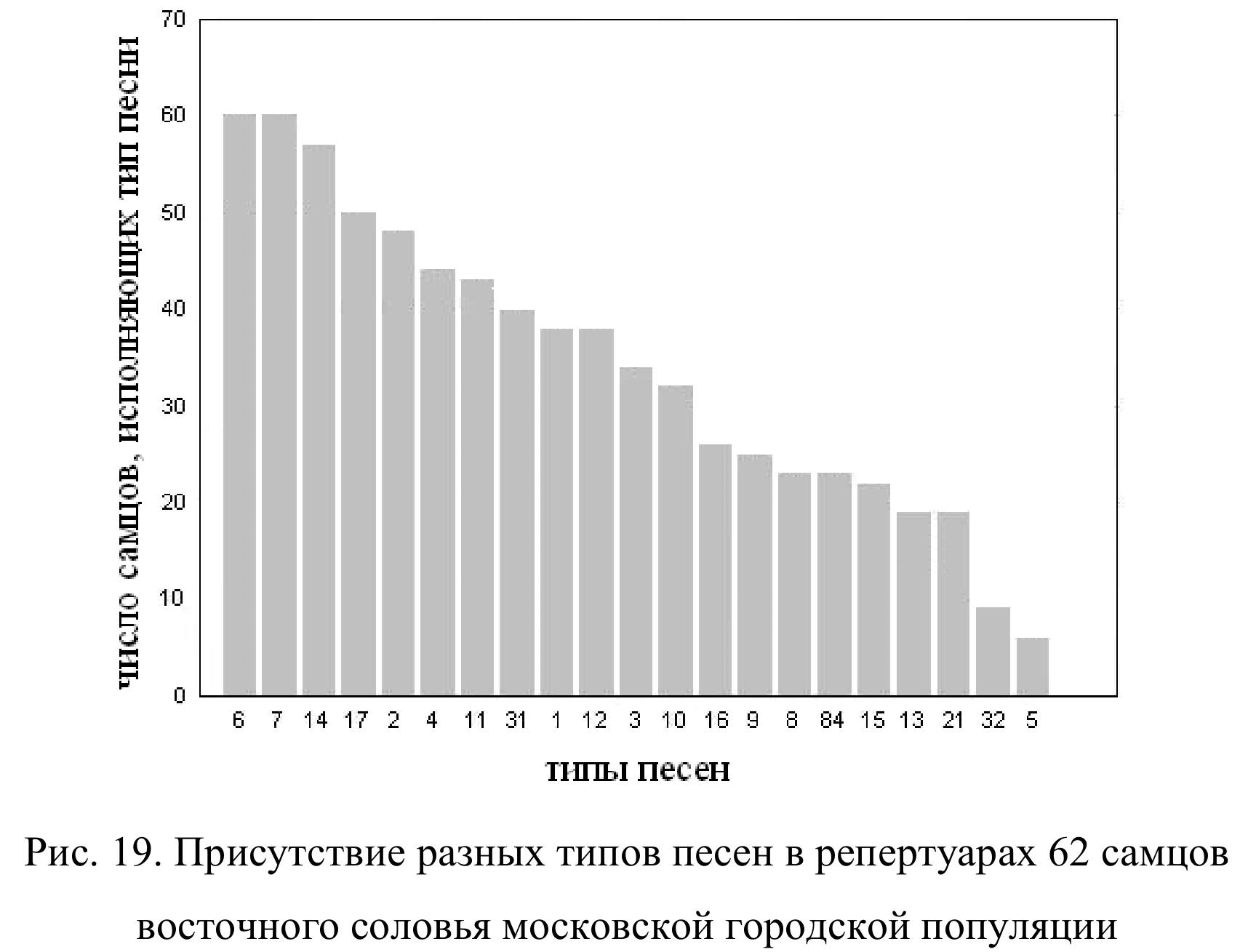
В этой главе мы будем анализировать синтаксическую организацию вокальных сессий восточного соловья на примере московской городской популяции, изученной нами наиболее подробно. В анализ мы включаем данные по 62 самцам, записанным в 2010 и 2011 гг. Размер их индивидуальных репертуаров варьирует от 4 до 21 типов песен. Медиана равна 11, квартили – 7 и 11. В общей сложности во всех 62 репертуарах мы нашли 44 типа песен. Из них 23 типа песен отмечены только у одного соловья и 21 тип песен исполняли как минимум 2 самца. Но при этом некоторые типы песен присутствуют в репертуарах многих или почти всех соловьев Москвы (рис. 19). Например, типы песен 6, 7, 14 и 17 отмечены у 50 – 60 (81.6 — 96.7 %) самцов. Из 21 типа песен 12 отмечены в репертуарах более чем половины всех соловьев. Таким образом, степень сходства репертуаров в целом довольно высока.
На рис. 21 представлены результаты многомерного шкалирования, выполненного с использованием полной матрицы индексов Жаккара. Для сравнения представлены также данные по 2014 году. Индекс Жаккара, вычисленный при парном сравнении всех 62 репертуаров, варьирует от 0.1 до 1.0 (две пары самцов) и составляет в среднем 0.47, что свидетельствует о высокой степени общности. Точками и эллипсами разного цвета на рисунке обозначены самцы, записанные в северном, южном, западном и восточном секторе города. Как видно из рисунка, облака точек, соответствующие разным секторам, широко перекрываются, причем за оба периода наблюдений получен, в сущности, очень близкий результат. Размещение точек в пространстве двух координат совершенно не соответствует


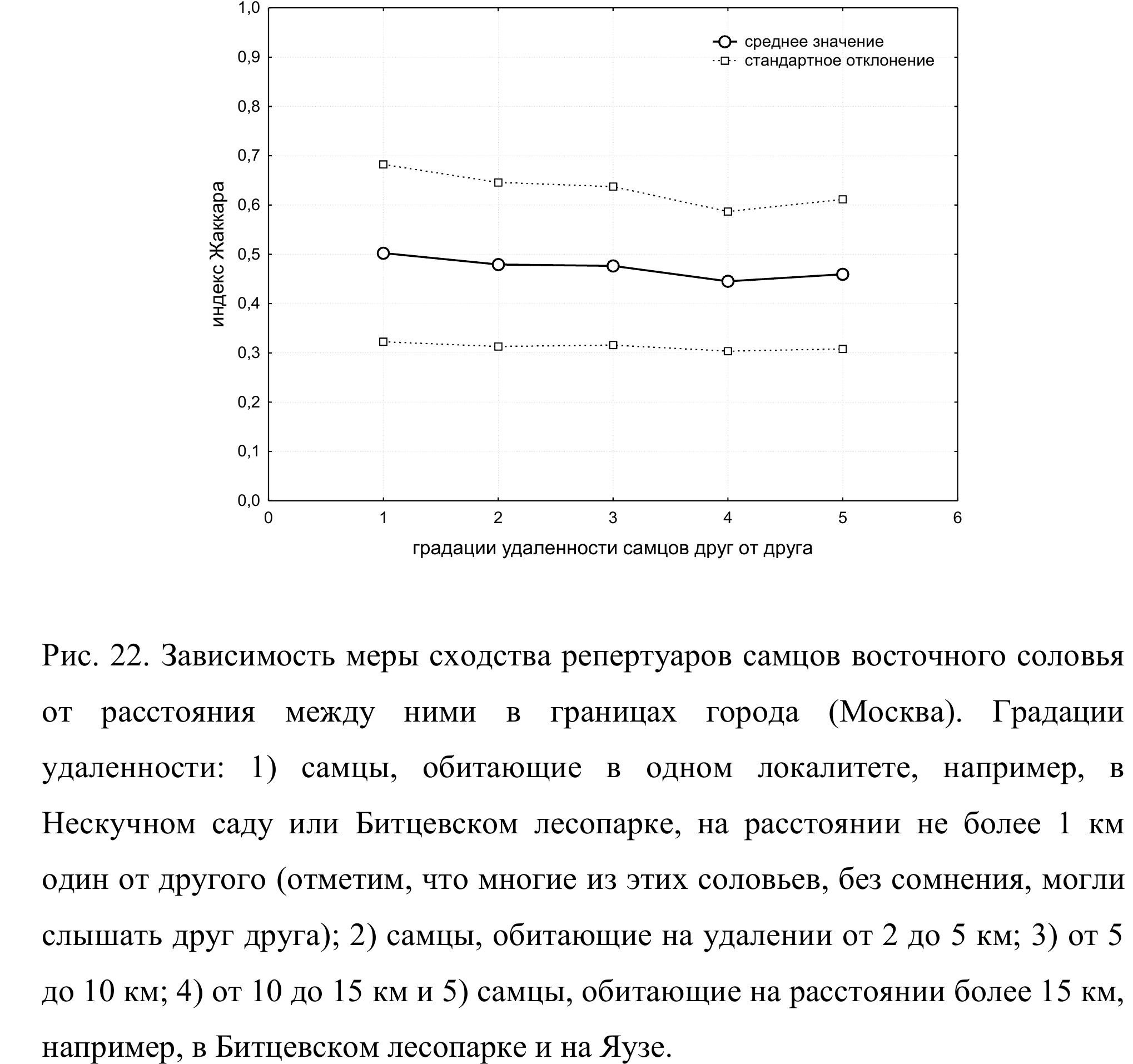
реальному распределению самцов по городским участкам. То есть, в пределах Москвы расстояние между двумя соловьями и мера сходства их репертуаров не связаны.
Количественную зависимость между мерой сходства репертуаров и расстоянием между самцами иллюстрирует также рис. 22. Мы выделили 5 степеней удаленности соловьев друг от друга: 1) самцы, обитающие в одном локалитете, например, в Нескучном саду или Битцевском лесопарке, на расстоянии не более 1 км один от другого (отметим, что многие из этих соловьев, без сомнения, могли слышать друг друга); 2) самцы, обитающие на удалении от 2 до 5 км; 3) от 5 до 10 км; 4) от 10 до 15 км и 5) самцы, обитающие на расстоянии более 15 км, например, в Битцевском лесопарке и на Яузе. По каждой градации из общей матрицы мы в случайном порядке выбрали по 30 – 40 значений индекса Жаккара. Поскольку в выборках присутствуют множественные измерения, средние и стандартные отклонения были посчитаны методом случайного генерирования повторных выборок в программе PAST. В этой же программе мы подсчитали достоверность различий между градациями с использованием метода непараметрической ANOVA с перестановками, устраняющими эффект множественных измерений (для одного самца коэффициент Жаккара был вычислен многократно), и коррекцией Бонферрони против множественных сравнений (в анализе участвуют 5 выборок).
Влияние фактора принадлежности к выборке в данном случае недостоверно, следовательно, степень сходства репертуаров соловьев, обитающих в границах МКАД, не зависит от расстояния между ними. На каком бы расстоянии друг от друга в границах МКАД не жили бы два соловья, с высокой долей вероятности можно ожидать, что в их репертуарах будет около 50 % общих типов песен. Это дает основание рассматривать городскую популяцию соловьев Москвы как гомогенную с биоакустической точки зрения и в дальнейшем анализировать ее как единое целое.
4.2 Совместная встречаемость разных типов песен в репертуарах
На рис. 23 представлены данные о совместной встречаемости разных типов песен в индивидуальных репертуарах московских соловьев. Как можно видеть, три кластера имеют высокую бутстреп поддержку (> 70%). Это означает, что включенные в них типы песен встречаются в одном репертуаре вместе чаще, чем по отдельности. Например, сочетание типов песен 6, 7, 14 и 17 найдено в репертуарах 46 из 62 самцов (74.2 %), сочетание 6, 7 и 14 – в репертуарах 57 самцов (91.9 %), а типы песен 6 и 7 найдены в репертуарах 60
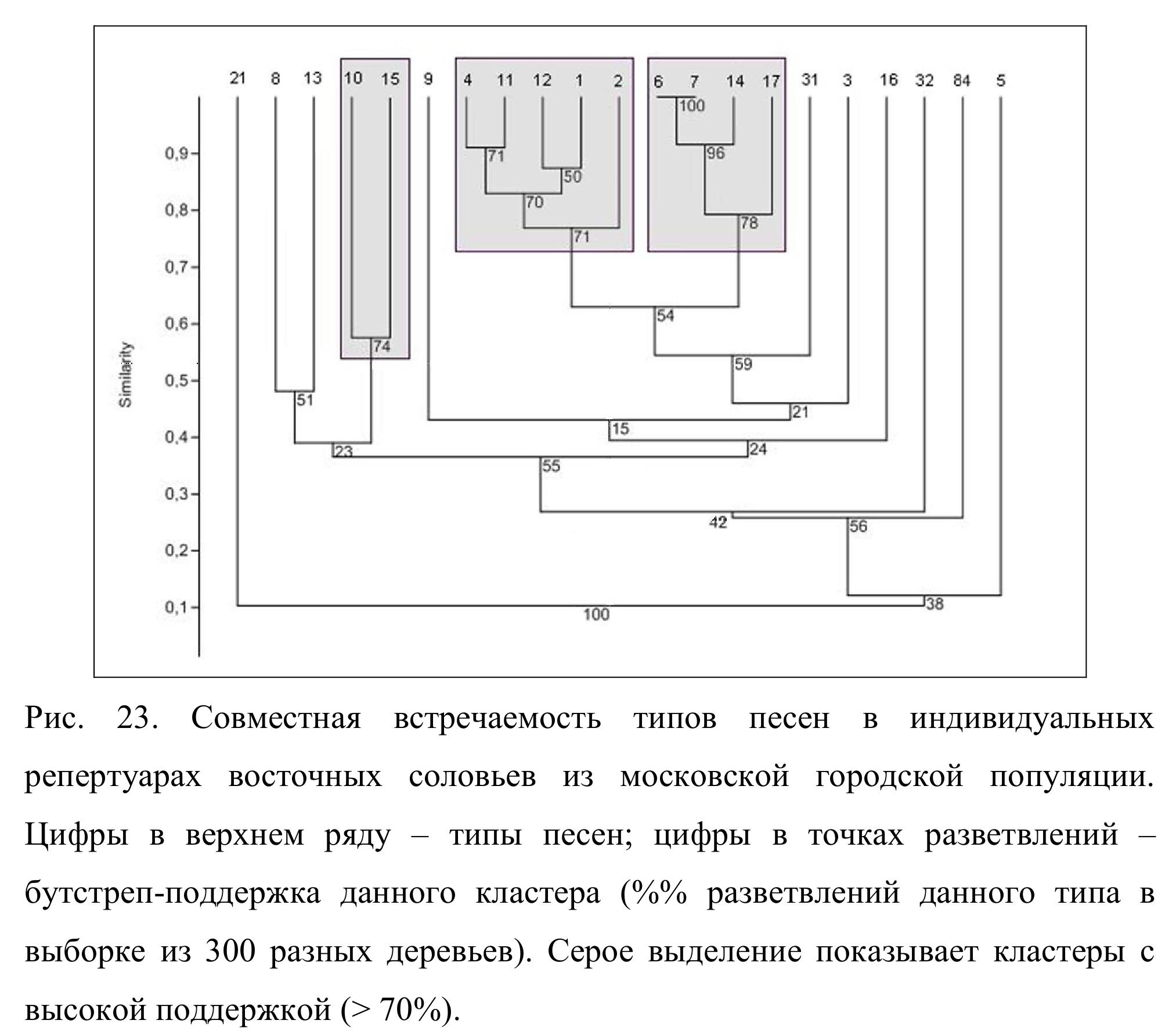
самцов (96.8 %). Столь же тесно ассоциированы между собой типы песен 1, 2, 11 и 12, а также 10 и 15. Кроме того, следует отметить высокую (59 %) бутстреп поддержку кластера, включающего типы песен 1, 2, 11, 12, 6, 7, 14, 17 и 20. Эти 9 типов песен найдены вместе более чем в половине всех репертуаров, и в общей сложности они составляют около 70 % всей вокальной продукции (общего числа песен на записанных нами фонограммах) московских соловьев. Таким образом, типы песен, наиболее широко распространенные в московской популяции восточного соловья, группируются в индивидуальных репертуарах неслучайным образом.
4.3 Переключения между типами песен по ходу вокальных сессий
Индивидуальные репертуары московских соловьев по объему различаются более чем втрое. Но даже и при самом бедном репертуаре (4 — 6 типов песен) исполнитель должен выбрать программу его презентации, то есть определенный порядок исполнения разных песен. Вопрос состоит в том, является ли выбор каждого очередного типа песни чисто случайным или же, например, зависит от того, какие типы песен были исполнены только что. Чтобы ответить на этот вопрос, мы определили порядок исполнения типов песен на всех 62 фонограммах, записанных нами в Москве в 2010-2011 году.
На рис. 24 показаны примеры «секвенирования» вокальных сессий 5 самцов восточного соловья из московской городской популяции. Номера здесь обозначают типы песен (в соответствии с рис. 4), а последовательность их исполнения читается сверху вниз (по столбцу) и слева направо (от одного столбца к следующему). Повторные исполнения одинаковых песен (подряд друг за другом) удалены. Для облегчения визуального восприятия рисунка деление непрерывной вокальной сессии на отрезки (столбцы), выполнено произвольно, но при строгом соблюдении истинного порядка исполнения. Разными вариантами заливки и обрамления выделены некоторые последовательности типов песен, устойчиво повторяющиеся по ходу пения. Одинаковое выделение использовано как для полных, так и для частичных (усеченных) последовательностей, включающих 2 и более песен.
На рис. 24 мы видим, что вокализация всех самцов содержит многократно повторяющиеся последовательности разных типов песен, исполняемые в определенном (постоянном) порядке. Причем часть из них, например последовательности [2→11→12→1→4], [17→14→6] и [13→4→9], присущи репертуарам разных самцов.


Отметим, что приверженность к стереотипным последовательностям выражена даже в тех случаях, когда в репертуарах нет полного соответствия типов песен. Сравним репертуары соловьев А и Б, записанных на Воробьевых горах (примерно в 800 м один от другого). Все типы песен из репертуара самца Б исполнял также самец А, репертуар которого включал также типы песен 3, 6, 8 и 17 (рис. 4). Коэффициент корреляции полных матриц переходов между типами песен в репертуарах этих самцов составил 0.55 при P<0.01. Связано это с тем, что значительная доля вокальной продукции обоих представлена полными и частичными вариантами последовательности [2→11→12→1], весьма популярной среди московских соловьев. Наряду с этим в репертуаре самца А также многократно повторялась и последовательность типов песен [13→4→8→9], тогда как у самца Б, не имеющего в репертуаре типа песен 8, ее замещала редуцированная последовательность [13→4→9]. Аналогичный пример дают полная последовательность [5→7→6→14] у самца А и ее редуцированный вариант [5→7→14] у самца Б, в репертуаре которого тип песен 6 отсутствует.
По результатам «секвенирования» мы составили итоговую матрицу переходов между разными типами песен по ходу вокальных сессий (Табл. 2). Продолжительность записанных нами фонограмм варьирует в широких пределах. Поэтому, чтобы в максимальной степени уравнять вклады отдельных соловьев, мы включили в матрицу данные по 42 самцам, фонограммы которых содержали от 90 до 100 песен (для более протяженных фонограмм все переходы после сотой песни в матрицу не включались).
Итоговая матрица содержит в общей сложности 3861 переход и в том числе 741 (19.2 %) переход между одинаковыми типами песен, расположенных в ячейках на диагонали. Обычно подряд повторяются только 2 песни (78.2 % всех повторений), намного реже – 3 (14.7 %) и лишь изредка – от 4 до 6 песен. Приверженность к серийному повторению одинаковых типов песен заметно варьирует у разных самцов и при этом по-разному проявляется для разных типов песен. Однако для 14 из 21 типа песни установлено, что они повторяются подряд достоверно чаще, чем это можно было бы ожидать по воле случая. Большинство из них по ходу вокальной сессии повторяются крайне редко или же не повторяются никогда, но зато некоторые повторяются практически всегда. Таким образом, в пении восточного соловья сочетаются непрерывная и периодическая вариативность. При исполнении очередной песни самец нередко повторяет предыдущую, однако все же чаще выбирает из своего репертуара новый вариант.
Поскольку в матрице суммированы данные по многим самцам, ее можно рассматривать как иллюстрацию универсальности переходов между типами песен. Распределение переходов по ячейкам матрицы выглядит крайне неравномерным и несимметрично относительно диагонали. Большинство переходов являются однонаправленными, что подтверждает способность восточного соловья запоминать порядок исполнения разных типов песен (Naguib, Kolb, 1992). Даже на итоговой матрице, суммирующей данные по многим исполнителям, отчетливо заметна векторная (однонаправленная) природа многих переходов. На индивидуальных матрицах она проявляется еще рельефнее. Например, фонограммы двух самцов содержали 174 и 138 прямых переходов (например, от типа песни А к типу песни Б) и, соответственно, лишь 14 и 11 обратных переходов (от Б к А). В обоих этих примерах различия между частотами прямых и обратных переходов высоко достоверны (двусторонний тест двух пропорций; P < 0.001). В целом же значения статистического критерия χ2, вычисленные для фонограмм, изображенных на рис. 24, свидетельствуют о том, что в каждом из пяти случаев очередность исполнения типов песен с высокой достоверностью (P < 0.001) отличается от случайных последовательностей 3.
Матрицы, построенные отдельно для самцов, обитающих в северном, западном и южном секторе города
(в восточном секторе записано слишком мало самцов), достоверно коррелируют друг с другом (R = 0.59–0.63; Mantel test, P < 0.01). Также достоверна корреляция матриц, построенных отдельно по результатам 2010 и 2011 гг. (R = 0.67; Mantel test, P < 0.05). Таким образом, последовательность исполнения многих типов песен выглядит очень сходной практически по всему городу.
3. В применении ко всей матрице критерий χ2 в данном случае дает лишь приблизительный результат; его статистически строгое использование затруднено, поскольку многие ячейки матрицы содержат нули или значения меньше 5. Поэтому мы посчитали достоверность для всех пар переходов с помощью точного критерия Фишера.
Примечательно, что наибольшее число переходов в итоговой матрице отмечается между типами песен, которые образуют кластеры на кладограмме (рис. 23). Наиболее яркий пример дают типы песен 2, 11, 12, 1 и 4 (рис. 4). Они не только часто встречаются совместно в индивидуальных репертуарах, но практически всегда исполняются именно в такой последовательности, причем многие самцы заключительный тип песен 4, как правило, исполняют дважды подряд. Как минимум один раз последовательность [2 → 11 → 12 → 1 → 4] была исполнена всеми 29 самцами, в репертуаре которых имелись эти типы песен. Число реверсий, то есть исполнений в обратном порядке, минимально. Важно отметить, что в позиции непосредственно перед типом песни 2 и в позиции сразу после типа песни 4 выбор вариантов типов песен выглядит более широким, нежели в промежутке между ними. Из этого можно заключить, что последовательность типов песен [2→11→12→1→4] представляет собой фиксированную вокальную композицию.
Типы песен 6, 7, 17 и 14 (см. рис. 4) присутствуют в индивидуальных репертуарах большинства московских соловьев и часто встречаются далеко за пределами города. Визуальный анализ сонограмм показывает, что эти типы песен тесно ассоциированы друг с другом в процессе пения. Рассмотрим подробнее очередность их исполнения. В общей сложности из 4 разных типов песен можно составить 24 разные последовательности. Мы подсчитали, сколько раз каждая из них повторяется во всем массиве данных, содержащем в общей сложности 5336 переходов между разными типами песен на фонограммах 62 самцов. Последовательность 6→7→1→14 птицы исполняли намного чаще всех прочих. Полностью она была исполнена 47 раз (данные о неполных исполнениях приведены на рис. 26). В полном виде она также найдена на 27 из 46 фонограмм, содержавших все 4 типа песен. Все прочие 23 возможные последовательности, которые можно составить из типов песен 6, 7, 17 и 14, были найдены намного более редкими. Из них девять не исполнялись ни разу, три были исполнены по 1 разу, еще три – по 2 раза, одна – 3 раза, две – 4 раза, одна – 5 раз и еще одна – 8 раз. Последовательность 17→14→6→7 при автоматическом подсчете в массиве данных была отмечена 29 раз. Однако в 18 случаях ей предшествовали типы песен 6 и 7. Иными словами, сама по себе, вне связи с типами песен 6 и 7, она была исполнена только 11 раз. Таким образом, приведенные подсчеты показывают, что последовательность типов песен 6→7→17→14 представляет собой стабильную композицию, даже вопреки тому, что матрица переходов между этими типами песен содержит немало реверсий между ними (табл. 2).
Что же касается типов песен 10 и 15, которые по данным кластеризации также часто встречаются вместе в составе индивидуальных репертуаров (рис. 23), то, насколько можно судить по итоговой матрице, они исполняются вместе друг с другом обычно в произвольном порядке и не намного чаще, чем вместе с прочими типами песен.
Мы полагаем, что приведенные выше данные свидетельствуют о том, что в репертуарах московских соловьев имеются фиксированные последовательности типов песен. По всей видимости, в памяти отдельных индивидов они сохраняются как единое целое и точно так же, как единое целое, передаются от одного исполнителя к другому по эстафете вокального обучения. Порядок исполнения составляющих их типов песен обычно в большой мере постоянен. Поэтому такие последовательности могут быть названы программами.
Рассмотрим теперь ситуацию с точки зрения теории вероятности. Если порядок типов песен случаен, то исполнение каждого из них можно рассматривать, как событие, не зависящее от исполнения всех прочих. Тогда, чтобы оценить ожидаемую вероятность исполнения сочетаний типов песни в определенном порядке (например, 1→2→3), следует перемножить их частоты и результат разделить на число вариантов выбора (три типа песен, например, можно исполнить шестью разными способами, то есть из них можно составить шесть разных последовательностей). Наблюдаемую частоту искомого сочетания песен находим, разделив фактическое число его исполнений на общее число всех сочетаний данного состава, имеющихся во всей последовательности песен. Это число определяем по методу «скользящей рамки» как N – (n — 1), где N – число песен на фонограмме, а n – число типов песен в искомом сочетании.
Для примера выполним вычисления для последовательности типов песен [2-11-12-1] в пении самца А (рис. 24). В полном составе эта последовательность прозвучала 7 раз. Всего на фонограмме, включающей 187 песен, имеется 184 разных последовательностей, состоящих из 4-х песен. Отсюда наблюдаемая частота исполнений последовательности [2-11-12-1] равна ~ 0.038. Ожидаемую вероятность этого события получаем, перемножив известные нам вероятности исполнения каждой из песен 1, 2, 11, 12 и разделив результат на 24 (число вариантов выбора для 4-х объектов).
Полученное таким образом значение вероятности (~ 0.000003) настолько невелико, что версию о возможности случайных повторений последовательности [2-11-12-1] приходится отбросить. Это верно даже для простейших (парных) сочетаний. Например, наблюдаемая частота исполнений последовательности песен [2-11] на фонограмме А (всего 17 исполнений) составляет ~ 0.09, тогда как ожидаемая вероятность ~ 0.007. Различия между этими пропорциями высоко достоверны (P < 0.001).
Таким образом, исполнение разных типов песен представляет собой процесс, детерминированный в очень высокой степени. Уже при наличии в составе репертуара 10 – 12 разных типов песен вероятность многократного случайного повторения их одинаковых последовательностей чрезвычайно мала. Для более обширных репертуаров, включающих, к примеру, свыше 15 типов песен, случайное повторение практически невозможно. Некоторые устойчивые связи между типами песен характерны для разных индивидуальных репертуаров, содержащих соответствующие типы песен. Например, программа [2-11-12-1] в полном виде и многократно исполнялась шестью самцами из московской популяции (пять из них представлены на рис. 24). Другая популярная программа включает типы песен [16-17-14-6]. Полностью она также отмечена в репертуаре шести самцов. После песни 16 обычно исполняется песня 17 (в 50 % всех случаев), затем с высокой вероятностью песня 14 (72.4 % случаев), затем чаще всего песня 6 (34.5 %). После песни 6 обычно звучит песня 7 (36.2 %), играющая роль своеобразного «переключателя». После песни 7 с равной и высокой вероятностью могли быть исполнены песни 17 и 14 или (с вдвое меньшей вероятностью) песня 6.
Программы [2→11→12→1→4] и [6→7→17→14] играют роль ядра московской вокальной культуры восточного соловья. Но у отдельных птиц имеются также и другие программы. Пение самцов с относительно небольшими репертуарами (6 – 10 типов песен) обычно имеет выраженную циклическую структуру и, скорее всего, представляет собой единую программу, исполняемую лишь с очень небольшим числом перестановок.
4.4 Секвенциональная организация вокальных сессий
Рассмотрим теперь секвенциональную организацию индивидуальных репертуаров с использованием методов теории информации. Мы построили информационные графы для 12 самцов московской популяции, для которых имелись длительные и непрерывные фонограммы, содержащие от 135 до 172 песен. На рис. 25 приведен итоговый граф, построенный по значениям медиан, вычисленных для всех соловьев по каждой позиции отдельно.

Первая точка графа соответствует максимальной энтропии последовательности (Hmax), принятой за 100 %. Ее величина зависит только от общего числа типов песен в данном репертуаре. Вторая точка показывает реальную энтропию, учитывающую численное соотношение разных типов песен во всей последовательности (в %% от Hmax). Наклон всех графов на этом первом отрезке относительно невелик, потому что частота исполнения разных типов песен из одного репертуара варьирует незначительно. Все последующие точки графа отражают изменения энтропии (неопределенности) в зависимости от числа исполненных песен. Иными словами, в каждой очередной точке мы оцениваем, как меняется мера неопределенности слушателя в ожидании новой песни, если он слышал одну, две, три и еще несколько предыдущих. Количественную оценку такого изменения вычисляем по формуле относительной неопределенности (Нi+1 – Hi) / Hmax. Эта величина дает нам меру предсказуемости конечного типа песен внутри сочетания определенного размера (двойки, тройки, четверки и т.д.), если известны все предыдущие типы (для двойки – 1, для тройки – 2 и т.д.).
Изменения предсказуемости отражены крутизной падения линии графа (величины энтропии). Наиболее глубокое падение мы видим между точками 2 и 3. Оно показывает значительное увеличение предсказуемости второго типа песни после того, как прозвучал первый. На следующих отрезках перепад значений по оси ординат уменьшается, однако, для большинства графов он выражен достаточно четко вплоть до 5-й или даже 6-й точки. Это означает, что, зная, например, четыре типа песни, исполненные подряд, мы можем предсказать пятый тип песни с большей вероятностью, чем если бы мы, зная только три последовательные типа, желали предсказать четвертый.
Мы попытались оценить достоверность изменений энтропии на разных отрезках графа с помощью непараметрического теста Вилкоксона для сопряженных пар наблюдений. Парой наблюдений в данном случае являются два значения энтропии одного и того же самца в двух соседних точках его графа. Несмотря на малый объем выборки (12 самцов) мы нашли достоверные различия между точками 2 и 3, затем 3 и 4, 4 и 5 и, наконец, 5 и 6 (при P < 0.05). Это означает, что энтропия (неопределенность) достоверно уменьшается всякий раз, когда мы переходим от одиночных типов песен к парам, затем к тройкам и к четверкам. Иными словами, вероятность правильного предсказания следующего типа песни является достоверно более высокой в тех случаях, если мы знаем: 1) один тип песни против полного незнания; 2) два типа песни против одного; 3) три типа песни против двух; 4) четыре типа песни против трех. Дальнейшее увеличение осведомленности на точность прогноза не влияет, то есть после прослушивания пяти типов песен, мы не сможем предсказать шестой с вероятностью более высокой, чем пятый тип песни после прослушивания четырех.
4.5 Поведенческие особенности исполнения программ
Программы представляют собой относительно устойчивые и стабильные вокальные конструкции. Это проявляется в целом ряде особенностей вокального поведения, которые можно наблюдать при их исполнении.
Например, программы исполняются «грамматически правильно» на любой скорости пения, которая зависит, главным образом, от протяженности пауз между отдельными песнями. Как уже отмечалось, в пении восточного соловья этот параметр варьирует в широких пределах. Средняя продолжительность пауз 2.6 с, но на максимальной скорости пения они сокращаются до 0.12 – 0.15 с. На такой скорости пение становится практически непрерывным (слитным). Любопытно, что в большинстве случаев при этом не только сохраняется структура отдельных песен, о чем уже было сказано, но и исполняются они в строгом соответствии со стереотипной программой. Хотя иногда можно наблюдать и полное объединение (слияние) соседних типов песен. Чаще всего это типы песен 11 и 12 (рис. 4). В репертуаре некоторых самцов они исполняются как раздельно, так и в виде объединенной стереотипной конструкции. Но во всех случаях их место в ряду типов песен строго соответствует программе.
Правильное исполнение программ имеет место и при очень медленном пении с продолжительными паузами (до 25 с) и не зависит от числа серийных повторений типов песен (число серийных повторений типа песни не влияет на его место в программе). В результате исполнение, к примеру, программы [2→11→12→1→4] растягивается иногда до 90 с, что почти вдвое превышает ее обычную продолжительность. Однако память безошибочно подсказывает соловью «грамматически правильный» тип песни на любой скорости исполнения: от 2 – 3 до 6 – 8 песен в минуту.
Еще одно проявление устойчивости программ мы видим в их сохранности вопреки весьма значительной изменчивости структуры составляющих их типов песен. О слиянии типов песен сказано выше. Также это относится и к распространенному у восточного соловья исполнению незавершенных (усеченных) песен. При этом всегда исполняется самое начало песни. Чаще всего это «запевка вместе с несколькими широкополосными звуками из центральной части песни. Автономного исполнения прочих компонентов песен (трелей всех типов и замыкающих элементов) не отмечено ни разу, т.е. соловей всегда берется за исполнение очередной песни с самого ее начала. Нередко несколько неполных (одинаковых или разных) песен исполняются подряд. Но порядок исполнения программы остается неизменным, даже если все составляющие ее песни исполняются в усеченном виде.
Но даже при полностью завершенном исполнении изменчивость типов песен на популяционном уровне весьма значительна. Один и тот же тип песен разные самцы нередко исполняют по-разному. Особенно сильно меняется число серийно повторяемых компонентов песни (нот или слогов). Примеры приведены на рис. 10. Крайние варианты изображенных там типов песен выглядят очень разными. Тем не менее, особенности структуры этих вокальных конструкций совсем не меняют их положения в последовательности типов песен. В любом исполнении, к примеру, тип песен 7 обычно следует за типом песен 6 и предшествует типу песен 17.
Еще одну интересную особенность вокального поведения восточного соловья иллюстрирует рис. 26. Там приведены данные о числе исполнений полных программ и их фрагментов во всем массиве наших данных (методика подсчета приведена выше). Серые прямоугольники показывают наблюдаемое число исполнений данной последовательности типов песен. При этом подсчитывались только неассоциированные, обособленные исполнения.
Например, при подсчете числа исполнений последовательности [7→17] мы не учитывали ее присутствие в более протяженных последовательностях [6→7→17→14], [6→7→17] и [7→17→14] (ассоциированные исполнения).
Черными линиями на рисунке показано ожидаемое число исполнений данной последовательности, вычисленное исходя из данных о вероятности исполнения песен данных типов (см. рис. 20). К сожалению, мы не можем подсчитать отдельно ожидаемое число только неассоциированных исполнений, а поэтому нельзя подсчитать обычным путем достоверность различий между наблюдаемыми и ожидаемыми числами.
Тем не менее, как нам кажется, данные, приведенные на рисунке, достаточно красноречиво свидетельствуют о том, что число полных и незавершенных (усеченных) последовательностей, начинающихся с типов песен 2 или 6, значительно превышает число последовательностей, которые начинались с других типов песен, например, с типа песен 11, 12, 7 и т.д.
Сравним очень длительную и сложную последовательность типов песен [2→11→12→1→4] и несопоставимо более короткую и простую последовательность [11→12]. В первом случае число наблюдаемых исполнений намного превышает число ожидаемых, тогда как во втором случае мы видим совершенно противоположное соотношение. Таким образом, в большинстве случаев соловьи начинают исполнять программы с начального типа песни. Вероятно, здесь можно провести четкую аналогию с манерой исполнения усеченных песен, которые птица всегда исполняет только с самого начала и никогда не начинает с середины песни.
