

Обыкновенный жулан (Lanius collurio) и ястребиная славка (Sylvia nisoria) часто встречаются в сообществах совместно. Вероятно, это обусловлено сходными требованиями к структуре местообитаний. Необходимым условием более или менее продолжительного сосуществования популяций в сообществе является как минимум частичное разобщение экологических ниш. Однако ястребиная славка и жулан в этом отношении изучены недостаточно. В публикациях можно встретить прямо противоположные взгляды. Так, большинство авторов утверждают, что конкурентные отношения и агрессивные контакты между данными видами отсутствуют, и это позволяет ястребиной славке, единственной из всего рода, обитать совместно с жуланом. Другие орнитологи, напротив, указывают на сильное перекрывание экологических ниш, приводящее к высокой степени конкуренции, что выражается и в прямом взаимодействии, и в нтерференции.
Поскольку пища выступает одним из важнейших ресурсов, зависимость от которого в природе не может быть снята, трофические отношения становятся основным параметром экологической дивергенции видов. Однако специфика состава пищи жулана и ястребиной славки в условиях симпатричного обитания практически не привлекала внимания исследователей. Опубликована только одна работа сравнительного характера, материал для которой собирался на территории Беларуси. Поэтому нашей целью было проведение аналогичного исследования в другой части ареала этих видов, где они также обитают совместно, с охватом как естественных, так и антропогенно-преобразованных стаций.
Задачи работы включали сбор и определение пищевых объектов ястребиной славки и жулана, определение специфики их птенцовой трофики в естественных и антропогенных ландшафтах, а также выявление наиболее стабильных видоспецифичных и изменчивых показателей питания птенцов изучаемых видов.
Материал и методы
Специфика птенцовой трофики ястребиной славки и жулана изучалась нами в 2004—2013 гг. на базе Рязанского государственного университета. Сбор материала по питанию гнездовых птенцов проводили на территории Рязанской области — как в природных стациях (Клепиковский район), так и в антропогенных (окрестности Рязани).
Данные по питанию птенцов собирали в течение гнездовых периодов изучаемых видов — с середины июня по середину июля. Кормовые объекты отбирали у птенцов методом шейных лигатур. Начиная с 2009 г. параллельно производили съемку процесса кормления птенцов при помощи видеокамеры. В статью вошли данные только по тем гнездам жулана и славки, которые располагались в непосредственной близости от гнезда другого вида — на расстоянии не более 50 м. Определено 167 кормовых объектов жулана и 349 — ястребиной славки.
Результаты
Состав пищи птенцов ястребиной славки и жулана на модельных участках показан в таблицах 1 и 2.
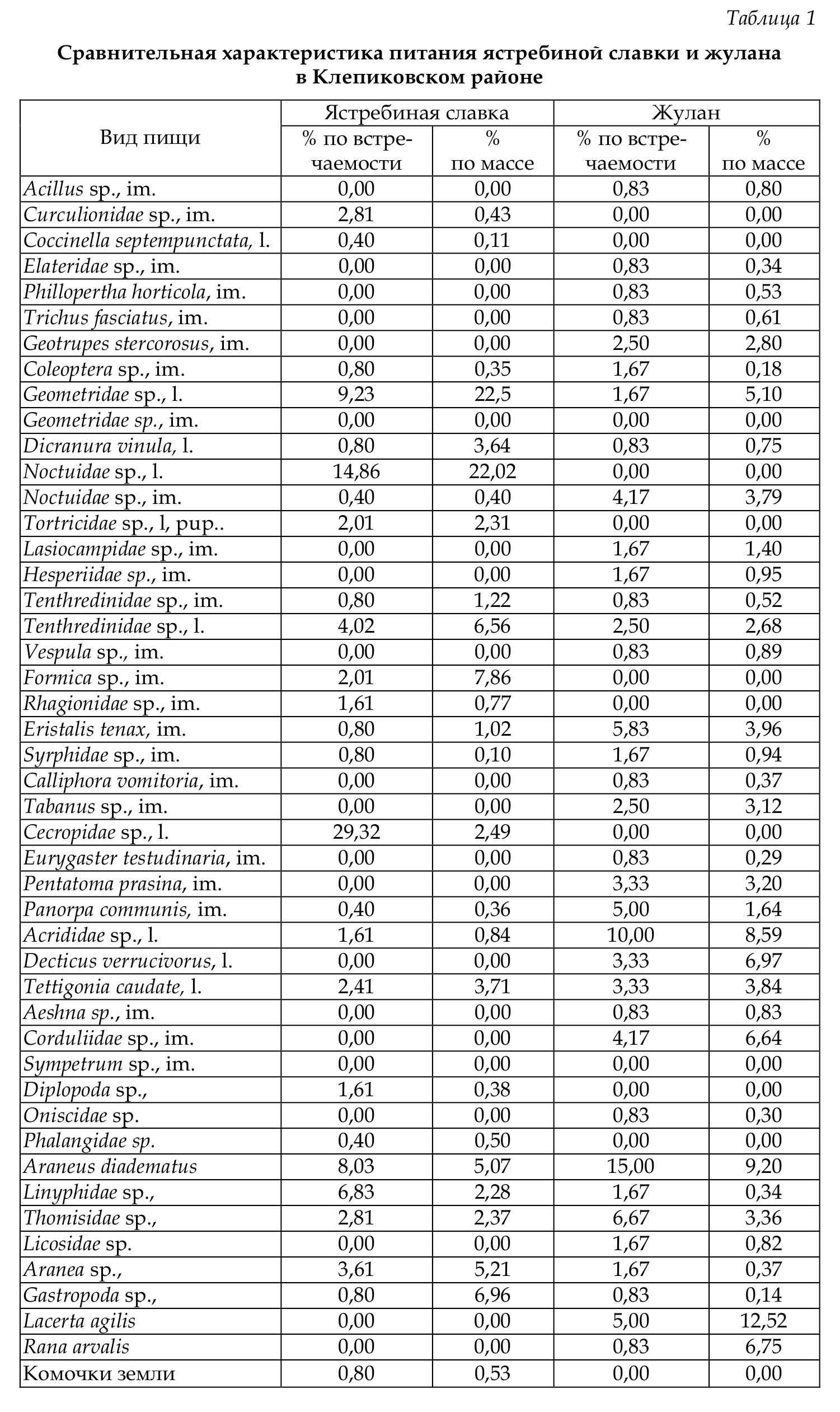
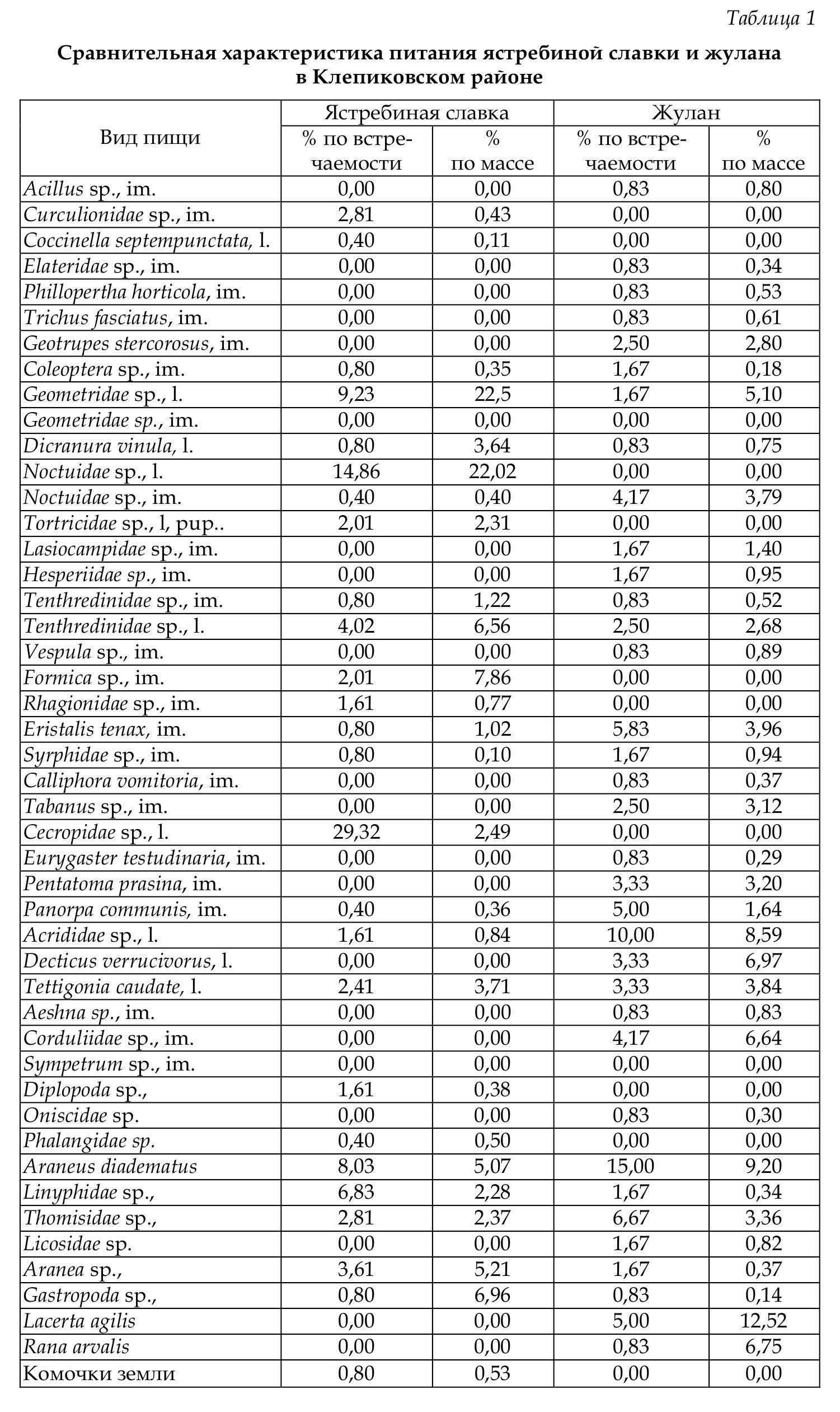
Сравнительный анализ полученных данных показал, что для обоих видов характерна значительная пластичность в питании. В каждой из исследованных нами стаций таксономический состав добычи птиц и массовая доля трофических групп существенно различались В Клепиковском районе в рационе птенцов ястребиной славки наиболее часто встречаются гусеницы и личинки пенниц. Однако масса последних невелика, они не играют важной роли в питании птенцов, несмотря на высокую встречаемость. По массе в рационе птенцов ястребиной славки преобладают гусеницы.
В пище птенцов жулана в Клепиковском районе также преобладают насекомые. В относительно большом количестве поедаются паукообразные, а также позвоночные (зарегистрирована ящерица прыткая и лягушка остромордая). В связи со сравнительно крупными размерами добываемых позвоночных их массовая доля в рационе довольно велика — 20,5 %.
По сравнению с другими птицами в питании птенцов жулана довольно высока доля жесткокрылых (6,3 %), которых обычно мелкие птицы съедают сами и редко приносят птенцам. При этом жуланы поедают таких крупных жуков с твердыми покровами, как жужелица, навозник, восковик перевязанный и др. Среди других беспозвоночных в пище этого вида преобладают крупные бабочки, мухи (пчеловидки, слепни и т.д.), разнокрылые стрекозы, клопы-щитники, кузнечики и кобылки.
В окрестностях Рязани по численности добытых славками особей гусеницы и личинки пенниц, преобладающие в Клепиковском районе, наряду с личинками пилильщиков, играют роль субдоминантов. Доминировали здесь имаго нехрущей, а также личинки и имаго кузнечиков рода Tettigonia. Этих крупных беспозвоночных славки начинали приносить уже 5—6-дневным птенцам, причем последние нередко проглатывали их с видимыми затруднениями и не с первой попытки. По мере роста птенцов потребление птицами хрущей и кузнечиков постоянно возрастало. В целом их суммарная массовая доля составила более 60 % рациона, на втором месте оказались ягоды (17,8 %).
В питании птенцов жулана в окрестностях Рязани также преобладали прямокрылые, на которых в сумме приходилось более четверти массы потребленной пищи. В несколько меньшем количестве поедались гусеницы совок, имаго нимфалид (в основном Nymphalis polychloros), еще реже жесткокрылые и арахниды. Позвоночные в рационе жулана здесь нами не были отмечены, несмотря на высокую их численность в кормовых стациях.
Очевидно, что таксономический состав пищи в значительной мере определяется структурой энтомофауны их кормовых микростаций, поэтому отмеченная нами пластичность питания обоих видов создает возможность тонкой подстройки к местным условиям, что, в свою очередь, оптимизирует кормодобывание и способствует снижению конкурентного пресса в каждом конкретном сообществе. При этом на фоне изменчивости таксономического состава пищи у каждого вида птиц должна проявляться присущая ему специфика других характеристик добычи. В результате даже в условиях симпатричного обитания пищевые спектры разных видов существенно отличаются, хотя в каждом случае наблюдается отчетливое влияние стациальных особенностей.
Статистическая обработка материала подтверждает это предположение. Так, коэффициент корреляции пищевых спектров ястребиной славки и жулана в Клепиковском районе по соотношению добытых птицами кормовых объектов составил – 0,34, а по их массовой доле в рационе + 0,02 при уровне значимости p < 0,01. В окрестностях Рязани соответствующие показатели составили + 0,36 и + 0,21 (p < 0,01). Таким образом, отсутствие как положительной, так и отрицательной скольконибудь заметной связи между данными свидетельствует о том, что в данном случае нет ни конкурентных отношений, ни специальных механизмов, направленных на их смягчение. Очевидно, что пищевые спектры обоих видов изменяются независимо друг от друга и определяются стациальными особенностями и видоспецифичным кормовым поведением.
Анализ полученных данных позволяет выявить некоторые наиболее стабильные и существенные особенности трофических адаптаций каждого из изученных видов. У ястребиной славки наряду с высокой пластичностью питания это в первую очередь преимущественное поедание открытоживущих беспозвоночных с мягкими покровами и потребление крупных объектов, добывание которых более энергетически выгодно для птиц.
У жулана основными особенностями рациона также можно назвать предпочтение открытоживущих беспозвоночных, что обусловлено особенностями кормового поведения вида. Жулан относится к типичным подстерегателям. Для него основным поисковым маневром при кормежке служит осматривание. Птица подкарауливает добычу, сидя на
присаде. Жулан атакует и схватывает жертву, используя дальний полет. Второй специфической чертой питания жулана является целенаправленное добывание объектов наибольших размеров, даже при условии наличия твердых покровов или высокой подвижности добычи.
Таким образом, основные отличия рационов рассматриваемых видов определяются тактикой поиска пищи: у жулана это поджидание добычи на присаде, а у ястребиной славки — поиск в кустарниковом ярусе и высокотравье. В частности, это определяет значительную роль в пище жулана подвижных, хорошо летающих насекомых, а у ястребиной славки — преобладание малоподвижных. Так, в Клепиковском районе в рационе ястребиной славки на хорошо летающих насекомых приходится 6,83 % кормовых объектов, составляющих 11,73 % массы рациона. У жулана соответственно 29,17 и 26,52 %. В Рязани эти показатели оказались практически аналогичными. У ястребиной славки на летающих насекомых приходится 7,78 % добычи с массовой долей всего 3,41 %, у жулана 25,60 и 29,60 % соответственно.
Предпочтение ястребиной славкой и жуланом крупных пищевых объектов выступает существенным лимитирующим фактором их синантропизации, поскольку в населенных пунктах и их окрестностях наиболее уязвимыми перед деятельностью человека оказываются именно такие беспозвоночные.
Список литературы
1. Барановский А.В. Особенности питания птенцов ястребиной славки // Экология и эволюция животных : сборник научных трудов кафедры зоологии РГПУ / под ред. Н.В. Чельцова. Рязань, 2004.
2. Кныш Н.П. Биологические особенности сорокопута-жулана как фонового вида лесостепной полосы УССР : автореф. дис. … канд. биол. наук. Киев, 1987.
3. Красная книга Московской области / под ред. Т.И. Варлыгиной, В.А. Зубакина, Н.А. Соболева. М., 1998.
4. Красная книга Рязанской области / под ред. В.П. Иванчева, М.В. Казаковой. 2-е изд. Рязань, 2011.
5. Красная книга Рязанской области / под ред. В.П. Иванчева. Изд. 1-е. Рязань, 2001.
6. Мальчевский А.С. Гнездовая жизнь певчих птиц. Л., 1959.
7. Мальчевский А.С., Кадочников Н.П. Методика прижизненного изучения питания гнездовых птенцов насекомоядных птиц // Зоологический журнал. 1953. Т. 32, № 2. С. 277—282.
8. Немчинов М.Ю., Мелеховец А.М., Сенченя А.Н. Различия в питании птенцов как аспект разобщения экологических ниш сорокопута-жулана (Lanius collurio) и ястребиной славки (Sylvia nisoria) // Материалы VII Междунар. науч.-
практ. конф. «Актуальные проблемы экологии» Гродно, 26—28.10. 2010. Гродно, 2011. С. 91—92.
9. Преображенская Е.С. Экология воробьиных птиц Приветлужья. М., 1998.
10. Птицы Рязанской Мещёры / под ред. Е. И. Хлебосолова. Рязань, 2008.
11. Птушенко Е.С. Род Locustella Kaup, 1816 // Птицы Советского Союза. М. ; Л., 1954. Т. 6. С. 239—271.
12. Птушенко Е.С., Иноземцев А.А. Биология и хозяйственное значение птиц Московской области и сопредельных территорий. М., 1968.
13. Симкин Г.Н. Певчие птицы. М., 1990.
Евгений Сергеевич Иванов — д-р с.-х. наук, проф., Рязанский государственный университет им. С.А. Есенина.
E-mail: o.hodosevich@rsu.edu.ru
Антон Валерьевич Барановский — канд. биол. наук, доц., НОУ ВПО «Современный технический институт».
E-mail: oldvulpes@yandex.ru
Источник
Вестник Балтийского федерального университета им. И. Канта. 2014. Вып. 7. С. 17—24