

Семейство: Тетеревиные Tetraonidae
Род: Тетерева Lyrurus
Вид: Lyrurus tetrix (Linnaeus, C 1758)
Протоним: Tetrao Tetrix.
Синоним: Lyrurus tetrix, Lyrurus tetrix tetrix, Lyurus tetrix, Tetrao empetrum, Tetrao tetrix.
Немецкое название: Birkhuhn
Испанское название: Gallo lira común.
Казахское название: қарағай-ұлар, қарақұр, Құр, саудырлақ
Украинское название: Тетерук.
Другие названия: полевой тетерев, косач, обыкновенный тетерев, тетерев-косач, березовик, черныш.
Taxonomic Serial Number : 677538 | Global Biodiversity Information Facility : 2473682.
Список ПОДВИДОВ вида Тетерев Lyrurus tetrix (Linnaeus, C 1758)
‐ Североевропейский тетерев — Lyrurus tetrix tetrix (Linnaeus, C 1758)
‐ Степной тетерев — Lyrurus tetrix viridanus (Lorenz, T 1891)
‐ Байкальский тетерев — Lyrurus tetrix baikalensis (Lorenz, T 1911)
‐ Тяньшаньский тетерев — Lyrurus tetrix mongolicus (Lönnberg, E 1904)
‐ Уссурийский тетерев — Lyrurus tetrix ussuriensis (Kohts, AE 1911)
ОСНОВНОЕ ОПИСАНИЕ ПО: Тетерев — Lyrurus tetrix (Linnaeus, C 1758)
Син. (русск.): тетерев-косач, полевой тетерев.
Син. (лат.): Tetrao tetrix Linnaeus, 1758; Tetrao betulinus Scopoli, 1769.
Статус. Оседлый, в ряде районов ареала нерегулярно кочующий вид.
Окраска. Взрослый самец почти целиком чёрный, с фиолетово-голубым оттенком на голове, шее, пояснице, надхвостье и груди, с белыми «зеркалами» на крыльях и белым подхвостьем. На спине синевато-голубоватый отлив выражен слабо, как и на крыле. Не имеет отлива также пространство вокруг клюва, бока тела и брюхо, имеющее чёрный или чёрно-бурый цвет. Задняя часть брюха бурая с широкими белесыми вершинами волосовидных перьев. Подхвостье белое, иногда с чёрными пятнами по краям наружных опахал нижних кроющих перьев хвоста. Первостепенные маховые бурые, с белесыми стержнями и белыми пятнами в нижней половине 1–5-го первостепенных маховых. Эти пятна — продолжение белого «зеркала», занимающего большую часть второстепенных маховых перьев. Эти перья белые, с широкой чёрно-бурой предвершинной полосой, занимающей почти всю верхнюю половину пера. Полоса расширяется на самых внутренних второстепенных перьях, так что на 1–2 самых последних перьях белый цвет вообще отсутствует.
Белые пятна в нижней части внешнего опахала больших кроющих перьев на развёрнутом крыле образуют самостоятельную белую полосу. Такие же белые июля имеют перья крылышка. Нижние кроющие крыла в основном белые и создают белый подбой, хорошо видный у взлетающей птицы. Рулевые перья чёрные с синеватым отливом на вершинах, изредка с белыми пятнами в основаниях. Первый взрослый наряд самца отличается меньшей интенсивностью синего отлива и значительным развитием коричневой струйчатости на плечевых, второстепенных и внутренних первостепенных маховых перьях, а также на верхних кроющих хвоста. Эта струйчатость исчезает обычно во втором взрослом наряде. Взрослый самец в летнем наряде; развивающемся обычно на голове и шее и в основном у южных популяций, приобретает пёструю окраску этих частей тела и нередко сплошь белое горловое пятно.
Самка во взрослом наряде пёстрая, при общем господстве коричневого цвета и рисунка из поперечных серых и черноватых полос. На вершинах перьев верхней части тела обычно развит серовато-чёрный струйчатый рисунок, степень развития которого определяет большую или меньшую сероватость общей окраски верха тела. Верхние кроющие крыла — с рисунком из чередующихся поперечных широких черно-бурых и узких тёмно-жёлтых полос; вершины белые со струйчатым рисунком (у некоторых птиц этих белых вершин нет). Окраска маховых перьев, как у самцов, но со значительным развитием коричневого струйчатого или поперечно-полосатого рисунка на окрашенной части пера. Нижняя часть тела несёт более равномерный, чем верхняя, поперечно-полосатый рисунок, особенно на груди, где чередуются тёмно-жёлтые и бурые полосы. Сильно рассученные вершины перьев несут здесь белые полоски, которые, будучи почти незаметными, значительно осветляют общий тон окраски. У ряда особей в верхней части груди имеется почти сплошная желтовато-рыжая окраска, а на горле — беловатое пятно, часто испятнанное мелким бурым крапом.
Наблюдается также два типа окраски нижней части груди и брюха. В одном случае здесь преобладает белесый цвет, создаваемый широкими белыми вершинами с незначительным струйчатым рисунком, а центр брюха занимает чёрно-бурое пятно. Во втором случае низ тела имеет равномерный поперечно-полосатый рисунок, несколько более мелкий по центру брюха. Подхвостье всегда белое. Нижние кроющие хвоста с широкими белыми вершинами и бурыми поперечными полосами, отороченными по нижнему краю рыжими полосками. Рулевые перья с вершинными белыми полосами и бурым струйчатым рисунком на вершинах, ниже которых идёт обычное чередование широких чёрно-бурых и узких тёмно-жёлтых полос. Оперение плюсны белесое или светло-серое.
Самка в первом годовом наряде отличается более чётким поперечно-полосатым рисунком на наружных опахалах первостепенных и второстепенных маховых перьев.
Самец и самка в ювенальном наряде пёстрые, с преобладанием тёмно-бурых или чёрных, жёлто-коричневых и белых полос и пятен. Преобладает поперечно-полосатый рисунок, особенно чётко выраженный на зашейке, спине и груди. На горле и передней части шеи преобладает рисунок из мелких бурых пятнышек. На нижней части груди и брюхе поперечный рисунок более тонкий и правильный. Стержни грудных перьев белые и образуют нечёткую продольную штриховку. На большинстве перьев крыла поперечные полосы разрезаются продольной белой стержневой полосой, постепенно расширяющейся к вершине и заканчивающейся пятном характерной треугольной формы. Рулевые перья поперечно-полосатые, с треугольным белым пятном на вершине.
Пуховой наряд лимонно-жёлтый снизу, с несколько более тёмной окраской груди. Верх тела коричнево-жёлтый, с неясными продольными и поперечными черно-бурыми полосами. На темени коричневая шапочка с чёрным окаймлением, по бокам головы ряд чёрных полосок и пятен.
Существует мнение, что 2-летних самцов можно отличить от следующей возрастной категории по длине и степени загиба наружной пары рулевых перьев. Данные, полученные при обработке большой серии птиц коллекции ЗИН АН СССР, вроде бы показывают возможность выделить по этому признаку даже 4 возрастные группы: молодых, 2-летних, 3-летних и более старших самцов; но значительная индивидуальная изменчивость этих показателей делает их ненадёжным критерием для точного определения возраста.
Самцы:
длина тела 508–708,
размах крыльев 805–897,
крыло 257–296,
хвост 93–118 (по длине центральной пары рулевых перьев), разница в длине центральных и крайних рулевых перьев 84–144, длина клюва 14.4–19.4,
плюсна 41–49,
средний палец 42–51.
Самцы в возрасте до года:
длина тела 542–603,
размах крыльев 802–870,
крыло 245–277, хвост 94–117,
разница в длине рулевых перьев 45–113,
длина клюва 13.5–18.4,
плюсна 40–49,
средний палец 40–50.
Самки (взрослые и молодые):
длина тела 470–508,
размах крыльев 770–800,
крыло 219–254,
хвост 88–123,
разница в длине рулевых перьев 10–36,
длина клюва 12.3–18.0,
плюсна 37–45,
средний палец 39–45.
Сезонные изменения массы типичны для тетеревиных птиц, но точное её изменение по сезонам не прослежено.
Как правило, масса самцов весной составляет 1 100 — 1 250, а осенью 1 200 — 1 350, но как весной, так и осенью встречаются особи с массой 1 500 — 1 600.
Максимальная масса самца, достоверно известная на сегодняшний день, 1 800 (Couturier, Couturier, 1980).
Масса самок колеблется в пределах 750 — 1 100.
Данные по массе 774 особей (Финляндия) дают следующие средние величины: в сентябре — 900, в октябре — 970, в ноябре — 990, в декабре — 980 (Koskimies, 1958).
Тетерев — стайная птица, и вне периода размножения держится стаями, как смешанными, так и однополыми. Состав стай, видимо, достаточно постоянен в течение осени и зимы, но точных данных на этот счёт нет.
Спят птицы как на деревьях, так и на земле, на снегу и в подснежных камерах.
Враги, неблагоприятные факторы. Определённый урон наносят тетеревам различные крупные хищники, среди которых на первом месте стоят лисица, куница и ястреб-тетеревятник. Лисица нередко охотится на тетеревов зимой во время их ночёвок в подснежных камерах, а также, как и куница, ловит птенцов и молодых птиц летом и осенью. Тетеревятник также охотится на тетеревов в течение всего года, но особенно ранней осенью (за молодыми птицами) и зимой. Следует отметить, что влияние хищников на численность тетеревов ничтожно по сравнению с тем уроном, который те несут под воздействием антропогенных факторов.
Из последних наиболее губительны используемые в сельском хозяйстве ядохимикаты и гранулированные удобрения, факторы беспокойства (прежде всего в период собирания ягод и грибов), а также линии электропередач. О последние разбиваются сравнительно много тетеревов в периоды, когда их активность начинается в темноте или в сумерках, то есть в условиях плохой видимости (период токования и осень). При малой численности вида также очень губительна охота на токах, к сожалению до сих пор очень популярная и практикующаяся в ряде мест. Из естественных факторов особенно неблагоприятны, как и для остальных тетеревиных птиц, длительные похолодания с затяжными дождями в период массового вылупления птенцов, зимы с частыми сменами оттепелей и морозов, а на юге ареала также засухи.
Хозяйственное значение, охрана. Из-за постоянного колебания численности правильная эксплуатация этого важного охотничьего вида возможна только при хорошо поставленном контроле за состоянием и движением численности. Промысловое значение тетерева оставалось высоким вплоть до середины нашего века, но сейчас оно сохранилось в лучшем случае только для северо-востока европейской части СССР и ряда районов Сибири. Во всех остальных областях значение тетерева не выходит за рамки спортивной охоты. Эксплуатация его в охотхозяйствах перспективна, но связана со значительными трудностями из-за подвижности птиц и постоянных колебаний их численности. В таких ситуациях особенное значение приобретает регламентация охоты вплоть до полного её прекращения при депрессиях. Основное внимание при этом должно быть направлено на охрану токовищ, весенняя охота на которых возможна при соблюдении тех же условий, что и при весенней охоте на глухаря.
Для поддержания численности вида в любом районе важное значение имеет правильно созданная система заказников, куда бы прекращался доступ людей в период размножения для снятия с птиц пресса фактора беспокойства. Было выяснено, что однократное вспугивание выводка в течение 7–10 дней подряд приводит к гибели птенцов (Рыковский, 1959). Из биотехнических мероприятий рекомендуется зимняя подкормка, создание искусственных галечников и порхалищ (Никульцев, 1967), однако наиболее эффективной может оказаться подкормка самок концентрированными кормами непосредственно перед началом размножения.
Всесоюзные учёты добычи тетерева по годам давно уже не проводились, но судя по тому, что ежегодная добыча этих птиц составляет в Норвегии около 80 тысяч, а в Финляндии от 54 до 134 тысяч (Couturier, Couturier, 1980), то в нашей стране она может превышать 1 миллион.
Для целей интродукции или реинтродукции тетерев мало подходит в силу его большой подвижности.

В СССР северная граница проходит через верхнюю часть Кольского залива (Семёнов-Тян-Шанский, 1959), южную оконечность Умбозера, водораздел рек Поной и Пана, по долинам рек Варзуга и Стрельна, с. Тетрино, к востоку от Белого моря она идёт через низовья реки Несь, устье реки Мезень, долину реки Сулы и её устье (кол. ЗИН АН СССР), южнее устья реки Ерсы (Портенко, 1937), устье реки Усы, долину Б. Кочмес (Данилов, 1975), низовья рек Собь, Полуй, Пельвож (Добринский, 1965; Бойков, 1965). Восточнее Оби северная граница точно не прослежена, её положение на рис. 37 дано по расспросным сведениям и по материалам колл. ЗИН АН СССР. Отдельные известные точки: Катарамбские озера, бассейн Среднего Вилюя, село Энгердек в долине реки Мархи (Андреев, 1974), долина реки Лены по 64° (Ларионов и др., 1980), долины рек Маи и Юдомы (Перфильев, 1975), междуречье Лены и Амги (Воробьёв, 1963), северная оконечность Байкала (Малышев, 1960), верховья Зеи, долина и устье реки Аргунь (Росляков, 1971), устье реки Горин (Воробьёв, 1954).
От последней точки граница идёт на юг, захватывая долину реки Уссури, окрестности озера Ханка и горы Северной Кореи, но нигде не выходит к тихоокеанскому берегу. Распространение в Китае точно неизвестно, и на рис. 37 граница ареала обозначена по очень приблизительной схеме Ченга (Cheng, 1978). Южная граница вновь входит в пределы СССР в районе верховьев реки Аргунь, обходя с севера Борзинские степи, и уходит в Монголию, где захватывает истоки рек Керулен и Онон, хребет Хэнтей и район озера Хубсугул. В Хангае тетерев пока не найден, нами же встречен в хребте Хан-Хухэй (Потапов, 1985). Далее на запад южная граница идёт по хребту Танну-Ола, обходит с севера пустынно-высокогорный и таёжный северо-восточный Алтай, от Телецкого озера идёт через центральный Алтай к западной части Катунского хребта и от него уходит в пределы Китая, захватывая там верховья реки Бухтармы (Сушкин, 1938).
Юго-восточнее этой границы сплошного ареала отдельные изолированные популяции обитают по юго-восточному берегу Телецкого озера от устья Кыги по верховьям правых притоков Чулышмана (Шавля, Чульча, Каракем), в верховьях Башкауса и в бассейне Сумульты (Фолитарек, Дементьев, 1938; Дулькейт, 1975). К западу от Алтая сплошной ареал тетерева распадается на ряд изолированных участков, самый крупный из которых захватывает горы Тарбагатая и Саура и мелкосопочник Восточного Казахстана, а остальные два включают Джунгарский Алатау и северный Тянь-Шань. Граница же сплошного ареала от предгорий Алтая идёт на север, вдоль боров долины Оби, а потом на запад, по южной границе лесостепи, оставляя южнее ряд изолированных участков (Наурзум, Тусумские пески, горы Улу-Тау), приуроченных крупным массивам боров или тугайной прирусловой растительности. По реке Урал идёт вниз до села Мергеневского (Кузьмина, 1962), хотя граница здесь весьма подвижна и при депрессиях значительно отступает к северу. Далее граница следует через Бузулукский Бор, по долине Самары, по южной окраине Жигулей, спускается по правобережью Волги до северных районов Волгоградской обл. (Кириков, 1975), вновь поднимается к северу и через северные части Воронежской, Курской, Сумской и Черниговской областей идёт на запад, уходя на территорию Польши, где захватывая её юго-восточную и восточную части, возвращается обратно в пределы СССР (Калининградская область и Литовская ССР) и выходит к побережью Балтийского моря под 57° северной широты.
Границы ареала указаны довольно приблизительно, ибо они постоянно изменяются как в результате колебаний численности вида, так и под влиянием человека. Сельскохозяйственное освоение степной и лесостепной зон Европы, Казахстана и Сибири вызвали заметное сокращение ареала, особенно на Украине и в Предкавказье, где ещё 100 с лишним лет назад тетерев водился до берегов Чёрного и Азовского морей и до подножья Кавказского хребта. В то же время отмечено некоторое расширение ареала на Кольском полуострове (Семёнов-Тян-Шанский, 1959). Ещё в верхнем плейстоцене ареал тетерева охватывал всю Европу до северных районов Испании и Италии включительно.
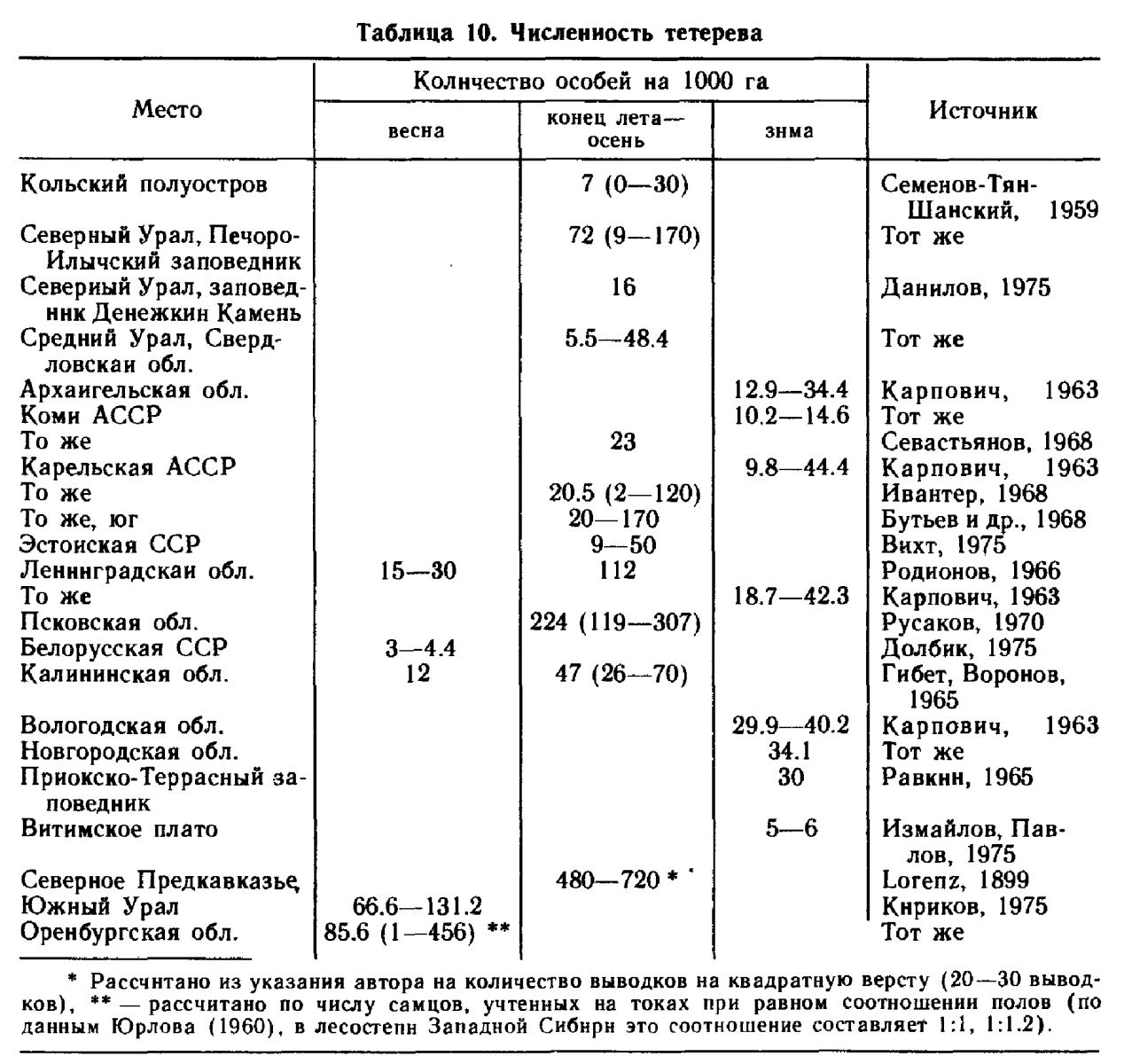
Численность. Наиболее высока в южной части ареала, особенно в лесостепи Восточной Европы и Сибири. Однако она нигде не бывает постоянной и испытывает колебания с амплитудой значительно большей, чем у остальных лесных тетеревиных птиц. Точное определение численности затрудняется большой подвижностью птиц и склонностью их к образованию скоплений. Это вызывает крайне неравномерное распределение их по территории, когда места с высокой концентрацией птиц сменяются местностью, где они совершенно отсутствуют. Конкретные данные о численности тетерева даны в табл. 10.
Другое исключение — обитание тетерева в степных вишенниках и ежевичниках, то есть в кустарниковых зарослях степей Украины. Предкавказья и западного Казахстана, наблюдавшееся в недавнем прошлом. В горах тетерев также отдаёт предпочтение окраинам сплошных лесов и разреженной древесной растительности, будучи наиболее обычным по нижней и верхней границам леса (как, например, на Алтае), либо же по одной из них. Так, в Альпах и большей части Карпат тетерев приурочен к верхней границе леса и нижней части субальпийского пояса, как и кавказский тетерев.
В качестве дополнительных кормов зимой используется большое количество других растений, среди которых наиболее часто встречаются ягоды шиповника, почки и побеги ивы, в Уссурийском крае даже преобладающие в рационе (Шульпин, 1936), почки осины, почки и серёжки ольхи. Весной в корме преобладают молодая зелень и прошлогодние ягоды, летом значительный процент составляют различные семена, а со второй половины лета начинают поедаться созревающие ягоды, полностью преобладающие в питании в августе — сентябре. Летом наиболее часто употребляются листья и ягоды брусники, черники и голубики, листья осины, соцветия ястребинки, цветочные почки сон-травы, листья манжетки, плоды черёмухи, листья клевера и семена осок. В сельскохозяйственных районах важной частью рациона становятся зерна культурных злаков, в особенности пшеницы и проса (Кириков, 1952; Ульянин, 1949; Юрлов, 1960).
В питании птенцов в первые дни преобладают насекомые, а нежная зелень, семена, плоды и ягоды употребляются в очень небольшом количестве. В Западной Европе птенцы в возрасте 12 дней поедают в основном насекомых — 76% суточного рациона, но уже в возрасте 2 месяцев этот процент падает до 6,25 (Briill, 1961). В западносибирской лесостепи, где питание птенцов изучено наиболее полно, они в первые дни жизни питаются только животной пищей. Растения, прежде всего листья клевера, начинают поедаться на 6-й день жизни. Доля растительного корма на 14-й день составляет 33,2% и с полуторамесячного возраста начинает преобладать. В лесостепи Западной Сибири молодые птицы в июле употребляют вдвое больше насекомых, чем взрослые, причём большая доля насекомых в рационе сохраняется до августа (Юрлов, 1960).
Следует отметить, что зимние корма сравнительно долго сохраняются в рационе птиц весной и довольно рано начинают употребляться осенью, нередко уже с лета, как в Барабинской лесостепи (Юрлов, 1960). Гастролиты в желудках содержатся постоянно, но в лесостепи их нередко заменяют твёрдые семена («костянки») различных растений, преобладающие над камешками в июле — ноябре и являющиеся единственными гастролитами в феврале (Юрлов, 1960).
Размеры токовища могут быть самыми различными, но в среднем токовище с 10–15 самцами занимает 0,3–0,5 га. При достаточной численности расстояние между токами в Ленинградской области может сокращаться до 1,1 км, а в лесостепи — до 900 м (Юрлов, 1960), но в среднем составляет 3–4 км. У изолированных популяций каждая из них имеет обычно одно токовище, отличающееся большой стабильностью, хотя здесь же могут быть и «запасные» токовища, где птицы токуют нерегулярно, а кроме того, некоторые самцы могут токовать в одиночку вне токовища. Одни и те же самцы в отдельных случаях могут поочерёдно токовать на разных токовищах (Юрлов, 1960).
Предположение, что каждое крупное токовище является репродуктивным центром данной микропопуляции, объединяющейся зимой в одну стаю, детальными наблюдениями не подтверждено (Юрлов, 1960; Потапов, 1985). Так, в Сибирской лесостепи территориальная структура популяций представляет собой участки от 12 до 40 км2, занимаемые отдельными микропопуляциями. Каждая из них состоит из 6–18 самцов и такого же количества самок. Самцы распределяются по нескольким токовищам числом от 3 до 7, по 1–8 самцов на каждом, и к каждому токовищу примыкает 1–6 гнездовых участков самок (осенью они дают 3–4 выводка). Основную роль в репродукции выполняет самое большое токовище, занимающее обычно центральное положение и служащее центром при формировании осенней стаи (Шинкин, 1966; Шило, 1972).
Весеннее оживление начинается в первых числах марта на юге ареала и во второй половине марта — в северной его части. Сначала это только пение отдельных самцов в стае на месте кормёжки («бормотанье» и «чуфыканье»), но постепенно интенсивность токования возрастает, и самцы начинают по утрам концентрироваться на постоянном или временном токовище. Токование поначалу идёт нерегулярно и любое ухудшение погоды надолго его прерывает. На льду озёр, где обычно нет никаких ориентиров для установления границ индивидуальных участков, драки между самцами происходят особенно часто.
С освобождением земли из-под снега самцы уже концентрируются на постоянных токовищах и самая активная фаза тока приходится на вторую половину апреля (юг) — первую половину мая (север). Длительность этого периода невелика, около 2 недель. Токование в это время начинается в полной темноте, почти за час до рассвета. Первыми начинают токовать старые самцы, прилетающие на токовище с соседних лесных опушек, хотя некоторые из них могут ночевать и на токовище. Они распределяются по своим токовым участкам и токование сразу же становится очень активным. Самки появляются тоже в темноте, но всегда позднее самцов, хотя иногда интервал между появлением самцов и самок составляет всего 15 минут (Юрлов, 1960). Самки рассаживаются сначала в непосредственной близости от токовища, издавая особые позывки «ак. . .ак. . .ак. . .», после чего слетают в центр тока. Средний по размерам ток посещают одновременно только несколько самок (1–4), причём далеко не каждый день. Само токование в течение утра идёт неравномерно — то усиливаясь, то ослабевая, то прекращаясь вовсе. Для Северо-Запада РСФСР характерны короткие, по 15–20 минут, перерывы, когда самцы смолкают и разом покидают токовище. Таких перерывов за одно утро может быть один или два. С восходом солнца интенсивность тока быстро падает, и через 1–1,5 часа токование прекращается.
Вечернее токование в разгар токовой активности столь же регулярно, как и утреннее, но никогда не достигает такой интенсивности, и спариваний на нём не происходит. Оно начинается с заходом солнца и продолжается до полной темноты, а на севере при коротких и светлых ночах — вплоть до утра. Поведение самцов на току строго территориально, каждый из них токует и спаривается только на своём участке, но постоянство последнего весьма относительно. В одних случаях на одном и том же токовище в центре его существуют из года в год одни и те же участки, занимаемые наиболее активными самцами, тогда как остальная территория каждый год делится каким-то количеством (тоже меняющимся) самцов, и каждый раз по-новому. В других случаях территориальная структура токовища каждый год меняется до неузнаваемости. Размеры участков тоже изменчивы. На постоянных токовищах при высокой численности птиц величина участка составляет всего 100 м (Hohn, 1953). В других случаях величина участка колеблется от 170 до 350 м2 в центре тока и до 4 000 м2 на его окраине. Самки стремятся приземлиться в местах с максимальной концентрацией самцов, то есть в центре токовища. Чем старше и активнее самец, тем ближе к центру токовища располагается его участок, тем выгоднее его положение для привлечения самок. Стремление самцов токовать в центре приводит с одной стороны, к тому, что центральные участки удерживаются самыми сильными самцами, а с другой стороны — к уменьшению размеров этих участков.
Иерархическая структура тетеревиного тока изучена ещё недостаточно полно. Считалось, что здесь существует трёхступенчатая иерархия: первую ступень составляют самцы в возрасте 3 лет и старше, занимающие самые центральные участки; вторую — 2-летние самцы, занимающие участки вокруг центральных, и третью — молодые самцы по окраинам тока (Koivisto, 1965). Другие исследования (Кириков, 1952; Юрлов, 1960) показали, однако, что довольно часто участки вокруг центра тока наряду с 2-летними самцами занимают молодые самцы. Среди последних наиболее активны птицы из самых ранних выводков предшествовавшего года. По другим данным, самцы старше 4 лет уже неспособны выдержать конкурентную борьбу в центре токовища и оттесняются в стороны (Vos, 1983).
Поэтому правильнее говорить о 2-ступенчатой иерархии: наиболее активные взрослые самцы, токующие в центре токовища, и все остальные. Действительно, на каждом нормальном токовище есть 1–3, а то и больше самцов, занимающих центральные участки, раньше начинающих и позже всего заканчивающих токование, наиболее активно токующих, на долю которых приходится около 98 % всех спариваний, происходящих на токовище (Koivisto, 1965; Vos, 1983; Потапов, 1985). Неоднократные высказывания о том, что спаривание тетеревов всегда происходит вне токовища (Сабанеев, 1876; Мензбир, 1902; Ульянин, 1949; Юрлов, 1960; Чельцов-Бебутов, 1965, и др.), ошибочны. Никто, по крайней мере до сих пор, не наблюдал непосредственно спаривания вне токовища. Все длительные наблюдения на токовищах в течение всего активного периода тока всегда устанавливали спаривания именно на самых центральных участках токовища (Koivisto, 1965; Kruijt et al., 1972; Vos, 1983; Потапов, 1985).
Одиночное токование тетеревов наблюдается обычно в районах с низкой численностью птиц, но оно бывает и там, где тетеревов много. Вполне возможно, что возникновение новых токовищ происходит именно на местах токования таких одиночек. Одиночное токование протекает по той же схеме, что и на коллективном токовище. Нередко такие самцы токуют в зоне непосредственной слышимости коллективного тока, иногда даже всего в нескольких сотнях метров от него. Сюда тоже прилетают самки, по 1, по 2, и самец спаривается несколько раз за сезон (наши наблюдения в Ленинградской области), но бывает, что самок около таких одиночных самцов не появляется вовсе. После того, как самки перестают посещать токовище, активность токования самцов постепенно затухает, но всё же продолжается на рассветах вплоть до начала — середины июня, окончательно прерываясь только с началом активной линьки. Самки, потерявшие кладку, могут вновь появляться на токовище и снова спариваться (Юрлов, 1960).
Брачные вокализации самцов можно слышать практически в течение всего года — и летом, и в середине зимы. Начиная с августа, «бормотанье» и «чуфыканье» самцов становятся слышны всё более часто, и максимальной активности осеннее токование достигает в конце сентября — начале октября. Но осенью самцы могут токовать где угодно, в том числе и на токовище. На последних в начале октября наблюдалось также территориальное поведение и даже драки (Птушенко, 1938). В середине зимы «бормотание» тетерева бывает слышно во время оттепелей, при тихой погоде. Совершенно особое явление — зимнее токование в Хэнтэе, описанное Козловой (1930). Ежедневно утром, перед кормёжкой, самцы прилетали на засыпанное снегом токовище и интенсивно токовали около часа. Токование продолжалось почти до конца января. Все добытые здесь птицы были старыми, самок вблизи токовавших самцов не наблюдалось.
Основная токовая поза самца, как и у белых куропаток, характеризуется горизонтальным положением вытянутой вперёд расширенной шеи и головы, которая даже слегка опущена. Хвост полностью раскрыт, поднят вертикально или слегка запрокидывается на спину. Резкий изгиб крайних рулевых перьев приводит к тому, что при максимальном, на 180°, раскрытии хвоста вершины перьев почти касаются земли (рис. 38).

Белые вершины нижних кроющих перьев хвоста выступают над короткими центральными рулевыми перьями, резко выделяясь на их чёрном фоне. Токовые прыжки очень характерны и в разгар тока совершаются постоянно, сопровождаясь специфичной вокализацией — «чуфыканьем». Во время таких прыжков птица может разворачиваться на 45–180°. При токовом взлёте птица с громким хлопаньем крыльев поднимается полого вверх и круто спускается вниз, делая 3–4 взмаха крыльями на взлёте и на спуске. Токовой полёт, обычно выполняемый в разгар токования, отличается от токового взлёта только тем, что достигнув высшей точки его, самец летит некоторое время горизонтальным полётом, облетая свой участок, и садится с громким хлопаньем крыльев. «Чуфыканье» — громкий шипящий звук из двух слогов «чуххх-ххы» с повышением тона в конце, длящийся от 0,9 до 1,1 секунды. Он издаётся в моменты наивысшего возбуждения, нередко предшествуя прыжку или взлёту, причём поза самца в этот момент вертикальная — шея и голова подняты вверх, клюв широко открыт, а перья на подбородке («борода») встают торчком.
Иногда чуфыканье звучит, как громкое односложное шипение «хисссс», издаваемое обычно во время токового полёта. Основной элемент брачного поведения — громкая своеобразная песня — «бормотание». Её максимальная слышимость около 3,5 км, а продолжительность от 2,2 до 4,2 секунд. В разгар токования перерывы между песнями, исполняемыми одним самцом, нередко не превышают 1 секунду. Песня состоит из мягких булькающих звуков, исполняемых как бы отдельными сериями. Сначала следуют несколько вступительных, с краткими паузами, звуков, птица как бы берёт разгон, а затем, раздув расширяющийся эзофагус, исполняет вторую часть песни из 15–18 булькающих звуков, содрогаясь всем телом от напряжения. Заключительная часть песни состоит из более отрывистых звуков такого же характера и примерно той же продолжительности, при этом все тело птицы содрогается от клюва до хвоста. Песня может исполняться как на земле, так и на дереве. Поёт самец всегда в основной токовой позе, вытянув расширенную шею вперёд параллельно земле. При конфронтациях самцов издаётся ещё один позыв — картавый хриплый вскрик типа «ку-карарарара». Особое явление — драки самок. Обычно это происходит, когда готовящаяся к спариванию самка видит другую в непосредственной близости. Нападающая самка раскрывает хвост на 160°, поднимает его под углом 45°, топорщит оперение и отставляет плечи в стороны так, чтобы было особенно заметно плечевое белое пятно.
Гнёзда устраиваются под прикрытием — под нависшими ветвями или у стволов деревьев, но бывают расположены и открыто — на лесных полянах или опушках в невысокой траве. Выстилка скудная, из сухой травы, листьев и нескольких перьев наседки. Глубина ямки 10-11 см, диаметр 23–28 см. В Казахстане гнёзда могут устраиваться даже в ковыльной степи. Есть сообщения о гнездовании самки на дереве, в 7 м над землёй, в старом гнезде какой-то врановой птицы (Witherby et al, 1947).
Окраска яиц от желтоватой до светло-охристой, с небольшим количеством мелких пятен и точек коричневого цвета, хотя этот крап бывает очень густой. Размеры яиц сильно варьируют. Длина колеблется от 46 до 58,4, ширина от 33,2 до 42,8. Средняя величина яиц из Альп (n = 21): 50,3 × 35,9, 47,6-52,1 × 33,8-37,2 (Couturier, Couturier, 1980). Масса свежих яиц (n = 128) 28,8–37,8, в среднем 33,9 (Юрлов, 1960),максимальная масса 40,94 (Couturier, Couturier, 1980). Наиболее обычное число яиц в кладке 7–9, в повторных кладках бывает 4–6. Максимальный размер кладки 13 яиц (Ивантер, 1968). Длительность насиживания 23–25 дней (Ульянин, 1949; Семёнов-Тян-Шанский, 1959; Юрлов, 1960). Срок, необходимый для откладки 1 яйца, колеблется от 1 до 2,2 суток, в среднем 1,5 суток. В процессе насиживания самка регулярно переворачивает яйца, в среднем 25 раз в сутки и оставляет гнездо для кормёжки от 1 до 4 раз, обычно 2–3 раза, затрачивая на кормёжку от 10 до 20 минут, иногда до 60–70 минут. Общая продолжительность отлучек составляет от 2,4 до 5,3 % всего времени насиживания, что влияет на продолжительность инкубации. Перед вылуплением птенцов самка не оставляет гнезда 19–20 часов (Семёнов-Тян-Шанский, 1959; Robel, 1969).
Вылупление птенцов происходит в течение нескольких часов. Масса новорождённого птенца 24–25 г (Юрлов, 1960; Федюшин, Долбик, 1967). Выводок покидает гнездо, и в первые сутки скорость его передвижения составляет около 10 м в час. Площадь перемещений выводка в середине июля (то есть с птенцами месячного возраста) составляет 16–17,5 га (Robel, 1969).
Сроки гнездования наиболее ранние в юго-западной части ареала, но в целом на всём его протяжении они изменяются незначительно. По всей южной части ареала время массового появления птенцов приходится на вторую декаду июня, в северной же части — на последнюю декаду июня — начало июля. Повторные кладки — нередкое явление, и пуховых птенцов можно встретить и в конце июля.
Основные места обитания выводков — окраины древесно-кустарниковых зарослей с обильными, иногда в рост человека, травами. В районе обитания выводка обязательно есть 2–3 места на солнцепёке, где птицы принимают солнечные и пылевые ванны — «порхалища». Птенцы начинают перепархивать в возрасте 10 дней. До месячного возраста они при опасности затаиваются, впоследствии же сначала затаиваются, а потом взлетают все вместе, разлетаясь в разные стороны.
Линяющие самцы всё лето держатся поодиночке или небольшими группами по 2–5 птиц, в самых глухих местах.
На деревьях также держится свободно и с большой ловкостью передвигается по самым тонким ветвям берёз, едва выдерживающим тяжесть птицы. Голосовые реакции довольно однообразны. Своеобразная «булькающая» песня самцов, издаваемая особенно часто и интенсивно во время токования, слышна и в остальные времена года, за исключением времени интенсивной линьки в июле и середины морозной зимы. При опасности самцы издают глухое отрывистое «гук.. . гук. . .» или серию звуков, издаваемых с различной частотой «тют-тют-тютт-тютт-тютт. . .тютт. . .тютт. . .тютт. . .тютт. . .» (Boback, Miiller-Schwarze, 1967). На тревожный позыв самки самец реагирует гортанным звуком «круаг. . .» или «крроаг», а иногда тревожный крик самца в стае звучит, как и во время токования, «ку-карр». Позывки самок в спокойном состоянии звучат, как тихие «как. . .как. . .гок». Учащение этих позывок уже означает тревогу. В случае непосредственной опасности, когда нет возможности скрыться, самцы и самки издают глухое шипение.
Размеры, чёрная окраска и своеобразная лировидная форма хвоста не позволяют спутать самца тетерева ни с одной другой птицей. В то же время пёстрая, с преобладанием серо-жёлтых тонов, самка очень похожа на глухарку и легко отличима от неё только в полете, благодаря белым «зеркалам» на крыльях.
В обычных условиях, при отсутствии массовых перемещений стая тетеревов ведёт сравнительно оседлый образ жизни. В Ленинградской обл. крупная стая держится обычно в течение всей зимы на ограниченной территории площадью примерно в 30 км2.
Тетерев значительно более чувствителен к морозам, чем глухарь или белая куропатка, и при морозах от —20°С и ниже стремится проводить в подснежной камере до 23 часов в сутки, затрачивая на кормёжку лишь 1 час, а то и меньше. В морозную погоду птицы покидают подснежные камеры с восходом солнца, совершая короткий, на 500–600 м, перелёт к месту кормёжки. Кормятся на берёзах, в зависимости от атмосферной температуры от 0,5 до 1,5 часов, после чего вновь летят на место ночёвки, которым бывает открытая поляна, просека, лесная дорога, зарастающая вырубка, верховое болото и тому подобного. Птицы постоянно меняют места ночёвок и, как правило, не зарываются повторно среди старых лунок. В конце зимы и ранней весной, с удлинением светового дня, птицы кормятся обычно дважды в сутки, утром и вечером, проводя середину дня снова под снегом или в поверхностной лунке. Во время оттепелей тетерев всегда отдыхает и ночует на деревьях. Чем ближе к весне и чем твёрже наст, тем больше времени птицы проводят на кормёжке, причём не только на деревьях, но и на снегу, расхаживая в поисках корма, как белая куропатка. Бюджет времени тетерева в середине зимы включает пребывание в подснежной камере — 15,6 часов, слабую активность (настороженная поза, чистка оперения и тому подобное) — 1,34 часа, дневной отдых на снегу или на дереве — 1,51 часа, кормёжку с передвижением по ветвям или по снегу — 4,91 часа, полёт — 0,05 часа.
Качество и количество зимнего корма определяются урожайностью того или иного растения, а также условиями зимы. При усилении морозов птицы стараются сократить время кормёжки за счёт преимущественного поедания концевых побегов и почек. Но если урожай серёжек хороший, то птицы питаются почти исключительно ими. Сухая масса суточного рациона в середине зимы составляет для самца 120 г и для самки 100 г. Серёжки, концевые побеги и почки встречаются в самых разных соотношениях.
Суточное количество твёрдых экскрементов при наиболее долгом, до 23 часов, пребывании в подснежной камере составляет 95–111 «колбасок». Масса экскрементов слепых кишек зависит от длины ночи и от атмосферной температуры и колеблется у самцов от 17,6 до 47,0 г (сухая масса), у самок от 7,1 до 37,4 г. Суточный бюджет энергии самца зимой: энергия, потребляемая с кормом — 2 472,2 кДж, экскреторная энергия 1 787,6 кДж, усвоенная энергия 735,6 кДж. Соответственно для самки эти параметры будут равны 2 101,4, 1 519 и 625 кДж (Потапов, 1982).
Размеры подснежной камеры 19-28×30-41 см, при высоте от 16 до 25 см. Толщина снежного потолка 7–11 см. Тетерев обычно не делает длинных подснежных ходов, проходя под снегом только 15–20 см. Но в отдельных случаях ходы могут достигать 4 м, с поворотом на середине на 90°. При раннем образовании наста птицы используют при зарывании в снег пробоины в насте на следах лосей — своего рода колодцы, пронизывающие всю толщину снега, через которые они также и покидают камеры.
Миграции. Тетерев отличается довольно подвижным образом жизни, и на большей части ареала постоянно происходят перемещения, особенно в годы депрессий или, наоборот, пиков численности. В то же время для большей части популяций известны и «оседлые» периоды их жизни, когда в течение ряда лет тетерева в определённой местности живут достаточно оседло.
Абсолютная оседлость свойственна небольшим изолированным популяциям Западной Европы, где и были определены размеры индивидуальных территорий, используемых отдельными особями в течение всего года. У взрослых самцов размер такой территории колеблется от 90–120 га в Альпах (Pauli, 1974) до 303-689 га в Шотландии и Голландии (Robel, 1969; Vos, 1983). Сезонные перемещения принимают временами значительные масштабы, но они не регулярны и связаны обычно с резким увеличением численности вида в каком-нибудь районе, равно как и со степенью обилия зимних кормов. Например, на среднем Амуре, в долине Сунгари и у озера Ханка большие массы тетеревов появлялись зимой, исчезая на лето (Шульпин, 1936).
Следующая линька — неполная летняя. Она наступает одновременно или несколько позже начала полной ежегодной линьки, стартующей у самцов в конце мая с выпаданием 1-го первостепенного махового пера. Летняя линька захватывает голову и шею, отчасти грудь, и бывает особенно хорошо выражена у самых южных популяций. На севере ареала масштабы её незначительны.
Максимальная выраженность летнего наряда у самцов — во второй половине июля — начале августа. В это время из-за бурной линьки маховых и рулевых перьев полёт птиц затруднён, они держатся очень скрытно, добывать их трудно, и они плохо представлены в коллекциях, отчего летний наряд долго оставался неизученным. Летняя линька у самок происходит в ещё более ограниченных масштабах, и в северной части ареала, кроме некоторого разреживания оперения, дальше не идёт. Полная годовая линька у самок начинается сразу же по окончании насиживания, в начале — середине июня, то есть несколько позже, чем у самцов.
Подробнее о сокращениях можно прочесть здесь.
Песня
…
Позывка
…
Электроманок
…
Литература
• Коблик Е.А. 2025. Фауна птиц стран Северной Евразии в границах бывшего СССР: Списки видов.
• IOC World Bird List v15.1 (Updated February 20, 2025) by Frank Gill, David Donsker & Pamela Rasmussen.
• Н.Н. Балацкий. Птицы Северной Евразии. Таксономический список: август 2023 г.
• «Птицы СССР. Курообразные. Журавлеобразные». т.2. Р.Л. Беме, Н.П. Грачев, Ю.А. Исаков, А.И. Кошелев, E.Н. Курочкин, Р.Л. Потапов, А.К. Рустамов, В.Е. Флинт. Ленинград. Издательство «Наука». 1987 г. стр. 233-248.
Сайты
• zmmu.msu.ru
• www.balatsky.ru
• www.worldbirdnames.org
• Avibase by Denis Lepage
• Zoonomen by Alan Peterson
• Taxonomy by John Boyd
• сайт Василия Вишневского
• www.xeno-canto.org
• www.featherbase.info, www.pbase.com, www.macaulaylibrary.org